- Administración de Parques Nacionales (SIB). (2025); Ficha: Piaya cayana; recuperado en junio 2025 de https://sib.gob.ar/especies/piaya-cayana?tab=avistajes
- Aleixo, A. (1997). Composition of mixed-species bird flocks and abundance of flocking species in a semideciduous forest of southeastern Brazil. Ararajuba, 5 (1):11–18. (enlace)
- Antelo, C. M. & Z. J. B. Fernández (2013). Las aves no passeriformes de Tucumàn, Argentina. Miscelánea, 132. Fundación Miguel Lillo, Argentina. (enlace)
- Avibase (s.f.). Ficha: Cuco ardilla común (Piaya cayana, Linnaeus 1766); recuperado en junio 2025 de https://avibase.bsc-eoc.org/species.jsp?avibaseid=F549E115E0ABD9B0
- Azpiroz, A., M. Mattalia, B. López-Lanús & L. L. Sosa (2013). Caracterización ornitológica y productiva del campo “Gral Ávalos” Monte Caseros, Corrientes, informe técnico de Aves Argentinas/AOP: diciembre 2010. En pp. 311-373: G. D. Marino, F. Miñarro, M. E. Zaccagnini y B. López-Lanús (eds.). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. (enlace)
- Babarskas, M., J. O. Veiga & F. C. Filiberto (1995). Inventario de Aves del Parque Nacional El Rey, Salta, Argentina. Monografía Especial L. O. L. A. No 6, 44 Págs. Buenos Aires.
- Babarskas, M., E. Haene & J. Pereira (2003). Aves de la Reserva Natural Otamendi. En Haene, E. y J. Pereira (Eds.). Fauna de Otamendi. Inventario de los animales vertebrados de la Reserva Natural Otamendi, Campana, Buenos Aires, Argentina. Temas de Naturaleza y Conservación. Monografía de aves argentinas, 3: 47-113. Aves Argentinas/AOP. Buenos Aires. (enlace)
- Baliño. J. (1984). Aves del Parque Nacional El Palmar, lista sistemática y otras contribuciones a su conocimiento. Rev. Mus. Arg. Cs. Nat. Zool., 13: 499-511
- Baliño, J. J. & F. J. Biole (1984). La avifauna de nuestros Parques Nacionales. Nuestras Aves, 5: 28-30. (enlace)
- Beltzer, A. H., P. A. Collins & M. A. Quiroga (2006). Atlas ornito-geográfico de la provincia de Entre Ríos. Serie Climax Nº 13. Ceride-Conicet. Santa Fe
- Bauni, V., R. E. Carini, S. Bogan, J. M. Meluso, T. N. Abad, M. Homberg & A. Giacchino (2017). Inventario de vertebrados del “Paisaje Protegido Delta Terra”, primera sección de islas del delta de Tigre, provincia de Buenos Aires, Argentina. Nótulas Faunísticas. Segunda Serie, 210: 1-11. (enlace)
- Belton, W. (1984). Birds of Rio Grande do Sul, Brazil. Part 1: Rheidae through Furnariidae. Bulletin of the American Museum of Natural History, 178: 369–636. (enlace)
- Berlepsch, H. (1887). Systematisches Verzeichniss der von Herrn Ricardo Rohde in Paraguay gesammelten Vögel, und Appendix Systematisches Verzeichniss der in der Republik Paraguay bisher beobachteten (enlace)
- Binford, L. C. (1989). A Distributional Survey of the Birds of the Mexican State of Oaxaca. Ornithological Monographs 43. American Ornithologists’ Union. Washington, DC, USA. (enlace)
- BirdLife International (2020). Species factsheet: Common Squirrel-cuckoo Piaya cayana. Downloaded from https://datazone.birdlife.org/species/factsheet/common-squirrel-cuckoo-piaya-cayana 19/06/2025
- Blake, E. R. & H. C. Hanson (1942). Notes on a collection of birds from Michoacan, Mexico. Field Museum of Natural History, Zoological Series, 22: 513-551. (enlace)
- Bodrati, A., Bodrati, G., Ferrari, C., Mérida, E. & E. Haene (1997). Notas sobre la avifauna del Baradero, provincia de Buenos Aires, Argentina. Nuestras aves, 37: 2-4. (enlace)
- Bodrati, A. & A. Pietrek (2000). Relevamiento de los recursos biológicos del Parque Provincial “Pampa del Indio” (Departamento Libertador Gral. San Martín, Chaco). AOP. Inf. Inéd. Buenos Aires.
- Bodrati, A., Mérida, E., Bodrati, G. & E. Sierra (2006a). Avifauna del talar de Vuelta de Obligado y sus ambientes contiguos. San Pedro, Pcia. de Buenos Aires, Argentina. E. Mérida & J. Athor J (Eds.) Talares bonaerenses y su conservación, 117-124. (enlace)
- Bodrati, A. (2006b). El árbol frutal de la selva misionera, Yabotikaba. Naturaleza y Conservación, 18: 10-15. (enlace)
- Bodrati, A., K. Cockle, J. M. Segovia, I. Roesler, J. I. Areta & E. Jordan (2010). La avifauna del Parque Provincial Cruce Caballero, Provincia de Misiones, Argentina. Cotinga, 32: 41-64. (enlace)
- Brandán, Z. J. & C. I. Navarro (2009). Lista actualizada de las aves de la provincia de Tucumán. Fundación Lillo, 12-55 (enlace)
- Burgos, F. G., J. L. Baldo & F. M. Cornell (2009). Lista de las aves de la Provincia de Jujuy, Argentina. Secretaría de Turismo y Cultura de Jujuy. S. S. de Jujuy. (enlace)
- Camperi, A. R. (1988). Notas sobre una colección de aves de los alrededores del Parque Nacional Baritú y del río Tarija, Salta. Hornero, 13: 21-33. (enlace)
- Camperi, A. R. (1990). Notas sobre una colección de aves del río Santa María, Salta, Argentina. Hornero, 13: 147-151 (enlace)
- Camperi, A. R. & C. A. Darrieu (2000). Estudio de una colección de aves del sur de la provincia de Misiones. Physis, Sec. C. 58 (134-135): 79-83.
- Capllonch, P. (1997). La avifauna de los bosques de Transición del Noroeste Argentino. Tesis doctoral, Facultad de Ciencias Naturales e Instituto M. Lillo, Universidad Nacional de Tucumán, 157 pp.
- Capllonch, P., Lobo, R., Ortiz, D. & R. Ovejero (2005). La avifauna de la selva de galería en el noreste de Corrientes, Argentina: Biodiversidad, patrones distribución y migración. Insugeo, Miscelánea, 14: 483-498. (enlace)
- Cardoso da Silva, J. M. (1994). Can avian distribution patterns in northern Argentina be related to gallery-forest expansion-retraction caused by Quaternary climatic changes? Auk, 111 (2): 495–499. (enlace)
- Carma, M. I. (2009). Relevamiento de aves de la provincia de Catamarca. Consejo Federal de Inversiones. 1º Ed., Buenos Aires. 500 pp. ISBN 978-987-510-081-7 (enlace)
- Carman, R. L. (1979). Piaya cayana en Buenos Aires. Hornero, 12 (1): 54-54. (enlace)
- Chatellenaz, M. L. (2004). La avifauna del bosque de Quebracho colorado y Urunday del noroeste de Corrientes, Argentina. Facena, 20: 3-12. (enlace)
- Chatellenaz, M. L. (2005). Aves del Valle del Río Paraná en la provincia del Chaco, Argentina: riqueza, historia natural y conservación. Miscelánea, 14: 528. (enlace)
- Chatellenaz, M. L., Cano, P. D., Saibene, C. & H. A. Ball (2010). Inventario de las aves del Parque Nacional Mburucuyá (Provincia de Corrientes, Argentina). Acta zoológica Lilloana: 139-160. (enlace)
- Chébez, J. C. (1993). Los manuscritos de William Henry Partridge. Aves Misioneras (VII). Nuestras Aves 28: 14–16. (enlace)
- Chébez, J. C. (1996). Fauna misionera, catálogo sistemático y zoogeográfico de los vertebrados de la provincia de Misiones (Argentina). Monografía No 5.; 320 pp. Ed. L.O.L.A. (Literature of Latin America). Buenos Aires. ISBN 950-9725-20-X
- Cherrie, G. K. (1892). A preliminary list of the birds of San Jose, Costa Rica. Auk, 9: 322-329. (enlace)
- Christiansen, M. B. & E. Pitter (1997). Species loss in a forest bird community near Lagoa Santa in southeastern Brazil. Biological Conservation, 80 (1): 23–32. https://doi.org/10.1016/S0006-3207(96)00073-0
- Cintra, R. & T. M. Sanaiotti (1990). Osteocephalus taurinus (treefrong): predation. Herpetological Review, 21: 59. (enlace)
- Cockle, K. L., Bodrati, A., Lammertink, M., Bonaparte, E. B., Ferreyra, C. & F. G. Di Sallo (2016). Predators of bird nests in the Atlantic forest of Argentina and Paraguay. The Wilson Journal of Ornithology, 128 (1): 120-131. (enlace)
- Coconier, E. G. (ed.) (2007). Las aves silvestres de Acambuco, provincia de Salta, Argentina. Relevamiento de un AICA prioritaria de la selva Pedemontana. Temas de Naturaleza y Conservación, 6: 1-127. Aves Argentinas/AOP, Buenos Aires (enlace)
- Contreras, J. R. (1979). Bird weights from northeastern Argentina. Bulletin of the British Ornithologists’ Club, 99 (1): 21–24. (enlace)
- Contreras, J. R. (1981). Lista preliminar de la avifauna correntina. I. No Passeriformes. Historia Natural, 2: 21-28.
- Contreras, J. R. (1987). Lista preliminar de la avifauna de la provincia de Formosa, República Argentina. Historia Natural, VII (4): 33-52.
- Contreras, J. R., L. M. Berry, A. O. Contreras, C. C. Bertonatti & E. E. Utges (1990). Atlas Ornitogeográfico de la Provincia del Chaco – República Argentina. Volume 1. No Passeriformes. Cuadernos Técnicos “Félix de Azara” 1.
- Contreras, J. R. (1993). Acerca de algunas especies de aves del extremo sudeste de la provincia de Formosa, República Argentina. Nótulas Faunísticas, 47: 1-8. (enlace)
- Contreras, J. R., F. Agnolin, Y. E. Davies, I. Godoy, A. Giacchino & E. Ríos (2014). Atlas ornito-geográfico de la provincia de Formosa, República Argentina. 1º Ed. Ciudad Autónoma de Buenos Aires: Fundación de Historia Natural Félix de Azara. 450 p. (enlace)
- Coria, O. R., Quiroga, O. B., Navarro, J. L., Heredia, J., Torres, R. & J. Lima (2021). Lista actualizada de las aves de Santiago del Estero, Argentina. Acta Zoológica Lilloana, 65 (1): 42–143. (enlace)
- Coria, O. R., Torres, R., De La Peña, M. R. & J. J. Lima (2023). Aves de Santiago del Estero. Ecología y Conservación. (1a Ed.). Buenos Aires: Fundación de Historia Natural Félix de Azara. (enlace)
- Cory, C. B. (1918). Catalogue of Birds of the Americas. Publications of the Field Museum of Natural History (Zoological Series) 197, 13 pt. 2 (2). 292 pp. (enlace)
- Darrieu, C. A., Camperi, A. R., Piloni, G. & N. R. Bogado (2013). Lista actualizada de las aves de la provincia de Buenos Aires. (1º Ed.) Fundación de Historia Natural» Félix de Azara». 56 pp. ISBN 978-987-29251-1-6. (enlace)
- de la Peña M. R. (1999). Aves Argentinas. Lista y Distribución. Ed. L.O.L.A. (Literature of Latin America). Monografía N.º 18; 244 pp. Buenos Aires. ISSN 0328-1620
- de la Peña, M. R. (2003). Nidificación simultánea de siete especies de aves (Passeriformes) en un bosque del centro de la provincia de Santa Fe. (enlace)
- de la Peña, M. R. (2006). Lista y distribución de las aves de Santa Fe y Entre Ríos. Monografía (16) L.O.L.A. (Literature of Latin America), Buenos Aires. ISSN 0-328-1620
- de la Peña, M. R., Laene Silva, R., Capuccio, G. & L. M. Bonin (2009) Aves del Río Uruguay. Guía Ilustrada de Especies del Bajo Uruguay y el Embalse de Salto Grande. (CARU). Comisión Administradora del Río Uruguay.
- de la Peña, M. R. & S. A. Salvador (2010). Manual de la alimentación de las aves argentinas. UNL. (Digital) (enlace)
- de la Peña, M. R. (2011). Atlas ornitogeográfico de la provincia de Santa Fe. Serie Naturaleza y Conservación No 4. Pág. 1-500.
- de la Peña, M. R. & R. F. Tittarelli (2011) Guía de aves de La Pampa. Santa Rosa. Gobierno de La Pampa.
- de la Peña, M. R. (2011). Atlas ornitogeográfico de la provincia de Santa Fe. Serie Naturaleza y Conservación No 4. 500 pp. ISBN: 978-987-27043-3-9
- de la Peña, M. R. (2012). Distribución y citas de aves de Entre Ríos. Santa Fe: Ediciones Biológica, Serie Naturaleza, Conservación y Sociedad, (6).
- de la Peña, M. R. (2013). Nidos y Reproducción de las aves argentinas. Serie Naturaleza, Conservación y Sociedad No 8. Ediciones Biológica. Santa Fe. (enlace)
- de la Peña, M. R. (2015). Aves Argentinas. Incluye nidos y huevos. 1º Ed. Santa Fe: Ediciones UNL; CABA: EUDEBA; Vol 1; 496 pp. ISBN 978-987-657-986-5
- de la Peña, M. R. (2016). Aves Argentinas: descripción, comportamiento, reproducción y distribución. Charadriidae a Trochilidae. Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie). Vol. 20 (1): 1-627.
- de la Peña, M. R. (2020). Aves Argentinas. Descripción, Comportamiento, Reproducción y Distribución (actualización). Cuculidae, Tytonidae, Strigidae, Nyctibiidae, Caprimulgidae, Apodidae, Trochilidae. Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie) N° 6: 1-348. ISSN 0325-3856
- del Hoyo, J. de. (2020). All the Birds of the World. Lynx Edicions, Barcelona. ISBN 978-84-16728-37-4
- De Schauensee, R. M. & Academy of Natural Sciences of Philadelphia (1964). The birds of Colombia, and adjacent areas of South and Central America. Narberth, Pa.: Livingston Pub. Co. 430 pp. ASIN: B0006BLJGW
- Dickey, D. R. & A. J. van Rossem (1938). The birds of El Salvador. Field Museum of Natural History Zoological Series, Vol. 23. Field Museum of Natural History, Chicago, Illinois. (enlace)
- Di Giacomo, A. G. & B. López- Lanús (2000). Nuevos aportes al conocimiento de la nidificación de algunas aves del noroeste argentino. Hornero, 15 (2): 131-134. (enlace)
- Di Giacomo, A. G. (2005). Aves de la Reserva El Bagual. En Di Giacomo, A. G. y S. F. Krapovickas (Eds.). Historia natural y paisaje de la Reserva El Bagual, provincia de Formosa, Argentina. Inventario de la fauna de vertebrados y de la flora vascular de un área del Chaco Húmedo. Temas de Naturaleza y Conservación 4: 201-465. Aves Argentinas/AOP. Buenos Aires. (enlace)
- Dubs, B. (1992). Birds of Southwestern Brazil. Catalogue and Guide to the Birds of the Pantanal of Mato Grosso and its Border Areas. Bertrona Verlag, Küstnacht, Suiza, pp. 1-164, pls. 1-58. (enlace)
- EcoRegistros (2025). Ficha: Piaya cayana; rcuperado en junio 2025 de https://www.ecoregistros.org/site/especie_lugares.php?id=333
- Echevarría, A. L., C. F. Marano & E. Fanjul (2008). Comunidades de aves del embalse Escaba y sus alrededores, Tucumán. Resumen de XII RAO: 166. San Martín de los Andes, Neuquén, Argentina.
- Erritzøe, J., C. F. Mann, F. P. Brammer & R. A. Fuller (2012). Cuckoos of the World. Christopher Helm, London, UK. 544 pp. ISBN 9781408142684
- Esteban, J. G. (1953). Nuevas localidades para aves argentinas. Acta Zool. Lilloana, 13: 349-362.
- Fandiño, B., Fernández, J. M., Thomann, M. L., Cajade R. & A. B. Hernando (2017). Comunidades de aves de bosques y pastizales en los afloramientos rocosos aislados del Paraje Tres Cerros, Corrientes, Argentina. (enlace)
- ffrench, R. (1991). A guide to the birds of Trinidad and Tobago. Second edition. Comstock Publishing Associates, Cornell University Press, Ithaca, New York. 524 pp. ISBN 0-7136-6759-1 (enlace)
- Fiora, A. (1933). El peso de las aves. Hornero, 5 (2): 174-188. (enlace)
- Fitzgerald, J., T. S. Schulenberg & G. F. Seeholzer (2020). Squirrel Cuckoo (Piaya cayana), version 1.0. In Birds of the World (T. S. Schulenberg, Editor). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.squcuc1.01
- Foster, M. S. (2007) The potential of fruit trees to enhance converted habitats for migrating birds in southern Mexico. Bird Conservation International, 17: 45–61. https://doi.org/10.1017/S0959270906000554
- Fraga, R. M. (2001). The avifauna of Estancia San Juan Poriahú, Iberá Marshes, Argentina: checklist and some natural history notes. Cotinga, 16: 81-86. (enlace)
- Friedmann, H. (1948). Birds collected by the National Geographic Society’s expeditions to northern Brazil and southern Venezuela. Proceedings of the United States National Museum, 97: 373–570. (enlace)
- Gasparri, B., Del Río, D., Earnshaw, A., Henschke, C., Bryant, G., Cristaldo, L., … & G. Suárez (2018). Lista comentada de las aves del Parque Natural Municipal Ribera Norte, San Isidro, Provincia de Buenos Aires, Argentina. Nótulas Faunísticas, 234: 1-43. (enlace)
- Gibbs, J. P. (1991). Avian nest predation in tropical wet forest: an experimental study. Oikos, 60: 155-161. (enlace)
- Gibson, E. (1919). Further Ornithological Notes from the Neighbourhood of Cape San Antonio, Province of Buenos Ayres. Part II. Trochilidae-Plataeide (enlace)
- Giraudo, A. R., Baldo, J. L. & R. R. Abramson (1993). Aves observadas en el sudeste, centro y este de Misiones (República Argentina), con la mención de especies nuevas o poco conocidas para la provincia. Nótulas Faunísticas, 49: 1-13. (enlace)
- Giraudo, A. R., Bortoluzzi, A. & V. Arzamendia (2006). Vertebrados tetrápodos de la Reserva y Sitio Ramsar “Esteros del Iberá” (Corrientes, Argentina): Análisis de su composición y nuevos registros para especies amenazadas. Natura Neotropicalis, 37 (1-2): 1-20. (enlace)
- Gomes, A.L.S., Marceliano, M.L.V. & M. A. Gonçalves Jardim (2008). Consumo dos frutos de Miconia ciliata (Rich.) DC. (Melastomataceae) por aves na Amazônia Oriental. Revista Brasileira de Ornitologia, 16 (4): 383–386. (enlace)
- Grant, C. H. B. (1911). List of Birds Collected in Argentina, Paraguay, Bolivia and Southern Brazil, with Field Notes. Part I. Passeres. Ibis, (9) 5 (17): 80-137; Part II. Picariae-Anatidae. (enlace) Ibis, (9) 5 (18): 317-350 (enlace); Part III. Columbidae-Rheidae. Ibis, (9) 5 (19): 459-478 (enlace)
- Griscom, L. & J. C. Greenway (1941). Birds of Lower Amazonia. Bulletin of the Museum of Comparative Zoology 88:83–344. (enlace)
- Gyldenstolpe, N. (1945). The bird fauna of Rio Jurua in Western Brazil. Kungliga Svenska Vetenskapsakademiens Handllingar Serie 3, 22 (3): 1–338. ISSN 0023-5377
- Haffer, J. (1975). Avifauna of Northwestern Colombia, South America. Bonner Zoologische Monographien 7. Zoologisches Forschungsinstitut und Museum Alexander Koenig, Bonn, Germany.
- Hartert, E. & S. Venturi (1909). Notes sur les oiseaux de la République Argentine. Novit. Zool., 16: 159-267. (enlace)
- Haverschmidt, F. & G. F. Mees (1967). Birds of Suriname.T. & A. Constable Ltd. Hopetoun Street. Printers to the University of Edimburg. Great Britain. (enlace)
- Hellmayr, C. B. (1929). A Contribution to the Ornithology of Northeastern Brazil. Publications of the Field Museum of Natural History, (Zoological Series) 255, Vol. 12 (18). Field Museum of Natural History, Chicago, USA (enlace)
- Herklots, G. A. C. (1961) The birds of Trinidad and Tobago. Collins, London, UK. 287 pp.
- Hilty, S. L. & W. L. Brown (1986). A Guide to the Birds of Colombia. Princeton University Press, Princeton, NJ, USA. ISBN 0-691-08371-1
- Hilty, S. L. (2003). Birds of Venezuela. Second edition. Princeton University Press, Princeton, New Jersey. 875 pp. ISBN 0-691-09250-8
- Howell, S. N. G. & S. Webb (1995). A Guide to the Birds of Mexico and Northern Central America. Oxford University Press, New York, New York, USA. (enlace)
- Junge, G. C. A. (1937). On Bonaparte’s types of the cuckoos belonging to the genus Piaya. Zool. Med. Rijksmuseum Nat. Hist. Leiden, 19: 183-185. (enlace)
- Junge, G. C. A. & G. F. Mees (1958). The avifauna of Trinidad and Tobago. Zoologische Verhandleingen number 37. (enlace)
- Klimaitis, J. F. & F. N. Moschione (1987). Aves de la Reserva Integral de Selva Marginal de Punta Lara y sus Alrededores. Dirección de Servicios Generales del Ministerio de Economía de la Provincia de Buenos Aires, Argentina.
- Klimaitis, J. F. (1987b). Primera cita del Tingazú Piaya cayana en el partido de Berisso (Bs. As.) y nuevo registro en Punta Lara, partido de Ensenada (Bs. As.). Garganchillo, 2: 7-8.
- Komar, O. & W. A. Thurber (2003). Predation on birds by a cuckoo (Cuculidae), mockingbird (Mimidae), and saltator (Cardinalidae). Wilson Bulletin, 115 (2): 205–208. (enlace)
- Kratter, A. W., T. S. Sillet, R. T. Cesser, J. P. O ́Neill, T. A. Parker & A. Castillo (1993). Avifauna of a Chaco. Physis, 16: 115-151
- Krauczuk, E. R. (2005). Aves do Inta-Campo Anexo ao Zaiman, e do Campus da Universidade Nacional de Misiones, Posadas, Misiones, Argentina. Atualidades Ornitológicas, 126: 17-39. (enlace)
- Krauczuk, E. R. (2008). Riqueza específica, abundancia y ambientes de las aves de Corpus Christi, San Ignacio, Misiones, Argentina. Lundiana, 9 (1): 21-31. (enlace)
- Laubmann, A. (1930). Vögel en Wissenschaftliche Ergebnisse der Deutschen Gran Chaco-Expedition. Verlag von Strecker und Schröder, Stuttgart
- Londoño, G. A. (2014). Parque Nacional del Manu, Cusco, Perú: Anidación de Aves en un Gradiente Altitudinal. Rapid Color Guide 514 versión 1. The Field Museum, Chicago, IL, USA. (enlace)
- López Lanús, B. (1997). Inventario de las aves del Parque Nacional «Río Pilcomayo» (Formosa, Argentina). Monografía especial Nº 4; 76 pp. Ed. L.O.L.A. (Literature of Latin America)., Buenos Aires, Argentina. ISSN: 0328-1620
- López-Lanús, B., A. S. Di Giacomo, A. Azpiroz, P. Haynes, A. Galimberti, A. Keyel, A. Ocampo, R. Güller, R. Moller Jensen, M. Mattalia, H. Cardoso, C. Giarduz, G. Papini & A. G. Di Giacomo (2013a). Inventario focal de fauna de las estancias La Higuera, María Concepción, La Sirena y Virocay en el sitio piloto Aguapey: Corrientes, Argentina. En pp. 179-223/277-293/307-319/331-347/490,494-506: G. D. Marino, F. Miñarro, M. E. Zaccagnini y B. López-Lanús (eds.). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. (enlace)
- López Lanús, B. & P. Mencía (2017). Diversidad, abundancia y estado de conservación de las aves de los riachos Monte Lindo y Pilagá en el este de la provincia de Formosa, Argentina. Nótulas Faunísticas (segunda serie), 228: 1-29. (enlace)
- Lucero, M. M. (1983). Lista y distribución de aves y mamíferos de la provincia de Tucumán. Miscelánea No 75. Fund. M. Lillo. Tucumán. 61 pp.
- Machado, R. B. & I. R. Lamas (1996). Avifauna associada a um reflorestamento de eucalipto no município de Antônio Dias, Minas Gerais. Ararajuba, 4 (1): 15–22.
- Malizia, L. R., Blendinger, P. G., Álvarez, M. E., Rivera, L. O., Politi, N. & G. Nicolossi (2005). Bird communities in Andean premontane forests of Northwestern Argentina. Ornitologia Neotropical, 16: 231–25. (enlace)
- Mancini, J. A. (2011). Inventario de las aves del Parque Escolar Rural Enrique Berduc. Entre Ríos.
- Marateo, G., H. Povedano & J. Alonso (2009). Inventario de las aves del Parque Nacional El Palmar, Argentina. Cotinga, 31: 47-60. (enlace)
- Martínez Gamba, R. (2014). Lista de aves del Parque Natural Municipal Monte Seguín, Provincia de Misiones, Argentina. Nótulas Faunísticas. Segunda Serie, 163: 1-10. (enlace)
- Mérida, E. & A. Bodrati (2006). Consideraciones sobre la conservación de los talares de barrancas del nordeste de Buenos Aires y descripción de las características de un relicto en Baradero. Pp.: 71-82, en : Mérida, E. y J.Athor (Editores). Talares bonaerenses y su conservación. Fundación de Historia Natural “Félix de Azara”. Buenos Aires. (enlace)
- Meyer de Schauensee, R. & W. H. Phelps (1978). A Guide to the Birds of Venezuela. Princeton University Press, Princeton, NJ, USA. (enlace)
- Miller, A. H. (1963) Seasonal activity and ecology of the avifauna of an American equatorial cloud forest. University of California Publications in Zoology, 66: 1–178.
- Mitchell, M. H. (1957). Observations on Birds of Southeastern Brazil. University of Toronto Press, Toronto, Ontario, Canada. 258 pp. ISBN 9780608163239
- Monroe, B. L., Jr. (1968). A Distributional Survey of the Birds of Honduras. Ornithological Monographs 7. American Ornithologists’ Union, Washington, DC, USA. (enlace)
- Moschione, F. & L. Bishels (2004). Listado de las aves del Parque Provincial Loro Hablador, provincia del Chaco. Informe Técnico. Proyecto Elé/DFS. Secretaría de Ambiente y Desarrollo Sustentable de la Nación.
- Moschione, F. N. & J. Segovia (2005). Listado de las aves de la Reserva Natural Las Lancitas, Santa Bárbara, Jujuy. (Informe Téc.) Proy. Elé/DFS.
- Moschione, F., Spitznagel, O. & M. Gonzalez (2014). Lista de Aves de Salta (Birds Checklist). Ministerio de Cultura y Turismo de Salta. Argentina. 47 p. (enlace)
- Narosky, T. & G. Di Giacomo (1993). Las Aves de la Provincia de Buenos Aires: Distribución y Estatus. Asociación Ornitológica del Plata, Vázquez Mazzini Editores & Literature of Latin America, Buenos Aires, Argentina. 127 pp. ISBN 950-99063-6-0
- Narosky, T. & L.M. Lozzia (1988) Aves de la Reserva El Bagual. Nuestras Aves, 15: 7-9. (enlace)
- Naumburg, E. M. B. (1930). The Birds of Mato Grosso, Brazil. A Report on the Birds Secured by the Roosevelt-Rondon Expedition. Bulletin of the American Museum of Natural History, 60: 1‒432. (enlace)
- Navarro S., A. G. (1992). Altitudinal distribution of birds in the Sierra Madre del Sur, Guerrero, Mexico. Cóndor, 94 (1): 29–39. (enlace)
- Navarro-Sigüenza, A.G. Y A. T. Peterson (2004). An alternative species taxonomy of the birds of Mexico. Biota Neotropica, 4 (2): 1–32. (enlace)
- Navarro, C. I. & N. L. Marigliano (2006). Avifauna otoño-invernal en un sector de bosque Chaqueño Serrano (Trancas, Tucumán, Argentina). Acta Zoológica Lilloana, 50 (1-2): 55-60.
- Nores, M., Yzurieta, D. & R. Miatello (1983). Lista y distribución de las aves de Córdoba, Argentina. Academia Nacional de Ciencias (Córdoba, Argentina). Tomo LVI: 1-119
- Nores, M. A. & D. Yzurieta (1979). Aves de Costas Marinas y de Ambientes Continentales, Nuevas para la Provincia de Córdoba. Hornero, 12 (1): 45-52. (enlace)
- Nores, M. & D. Yzurieta (1980). Aves de ambientes acuáticos de Córdoba y centro de Argentina. Sec. de Estado de Agricultura y Ganadería. Acad. Nac. Cienc. de Córdoba. Págs. 235
- Nores, M. A, D. Yzurieta & S. A. Salvador (1991). Lista y Distribución de las Aves de Santiago del Estero, Argentina. Bol. Acad. Nac. Cienc. Córdoba, 59: 157-196.
- Nores, M. (1994). Quaternary vegetational changes and bird differentiation in subtropical South America. Auk, 111 (2): 499-503. (enlace)
- Nores, M. (1996). Avifauna de la provincia de Córdoba. Fauna, 1: 255-337.
- Oniki, Y. & E. O. Willis (1999). Nest building and early incubation in Squirrel Cuckoos (Piaya cayana). Ararajuba, 7 (1): 23–25. (enlace)
- Ornelas, F. J., M. C. Arizmendi, L. M. Valdelamar, M. L. Navarijo & H. A. Berlanga (1993). Variability profiles for line transect bird censuses in a tropical dry forest in Mexico. Cóndor, 95 (2): 422–441. (enlace)
- Ortiz, D., Aráoz, R. & C. Alderete (2011a). Lista sistemática de las aves de la Reserva Experimental Horco Molle, provincia de Tucumán, Argentina. Nótulas Faunísticas, Segunda Serie, 62: 1-5. (enlace)
- Ortiz, D. & C. Ruiz (2011b). Migración y estacionalidad de aves en el arroyo Mista, Tucumán, Argentina. Nuestras Aves 56: 16-18. (enlace)
- Ortiz, D. & R. Aráoz (2014). Aves de la Sierra de San Javier. Ed. Universidad Nacional de Tucumán. Reserva Experimental Horco Molle. ISBN: 978-987-33-4376-6
- Ortiz, D., E. Martínez Paster, J. P. Juliá & P. Quiroga (2018). Lista de aves de la Reserva Privada San Pablo, provincia de Tucumán, Argentina. Nótulas Faunísticas. Segunda Serie, 246: 1-8. (enlace)
- Pagano, L. G. & E. Mérida (2009). Aves del Parque Costero del Sur. Pp. 200-244 en: Athor J (ed) Parque Costero del Sur. Naturaleza, conservación y patrimonio cultural. Fundación Félix de Azara, Buenos Aires, Argentina. (enlace)
- Pagano, L. G., E. A. Jordan, J. I. Areta, R .F. Jensen & I. Roesler (2012). Aves de la Reserva Natural Punta Lara. En 97-143: Roesler, I. y M.G. Agostini (eds). Inventario de los Vertebrados de la Reserva Natural Punta Lara, provincia de Buenos Aires, Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 8. Buenos Aires, Argentina. (enlace)
- Payne, R. B. & M. D. Sorenson (2005). The Cuckoos. Oxford University Press, Oxford, UK and New York, NY, USA. 618 pp. ISBN: 9780198502135
- Pereyra, J. A. (1950). Avifauna argentina (contribución a la ornitología). Hornero, 9 (2): 178–241. (enlace)
- Peters, J. L. (1940). Check-list of birds of the world. Volume 4. Museum of Comparative Zoology, Cambridge, Massachusetts. (enlace)
- Phelps, W. H. & W. H. Phelps Jr. (1958). Lista de las aves de Venezuela con su distribución. Volume 2, Part 1. No Passeriformes. Boletín de la Sociedad Venezolana de Ciencias Naturales, 19 (90): 1–317.
- Planqué, R., Vellinga, W. P., Pieterse, S., Jongsma J. & R. de By (Xeno-canto Foundation). Ficha: Cuco ardilla común (Piaya cayana, Linnaeus 1766); recuperado en junio 2025 de https://xeno-canto.org/species/piaya-cayana
- Pinto, O. M. O. (1938). Catalogo das Aves do Brasil e Lista dos Exemplares que as Representam no Museu Paulista. 1a parte. Aves não Passeriformes e Passeriformes não Oscines excluída a Fam. Tyrannidae e seguintes. Revista do Museu Paulista, 22: 1–566. (enlace)
- Pugnali, G. & P. Chamorro (2016). Lista de aves de la Reserva Costanera Sur. Aves Argentinas. Buenos Aires. (enlace)
- Pujalte, J. C., Reca, A. R., Balabusic, A., Canevari, P., Cusato, L. & V. P. Fleming (1995). Unidades ecológicas del Parque Nacional Río Pilcomayo. Anales de Parques Nacionales 16: 1-185. (enlace)
- Quiroga, O. B. (2019). Comentarios sobre nueve especies de aves de Santiago del Estero y Tucumán, Argentina. Nótulas Faunísticas. Segunda Serie, 274: 1-4. (enlace)
- Raw, A. (1997). Avian predation on individual Neotropical social wasps (Hymenoptera, Vespidae) outside their nests. Orn. Neotropical, 8 (1): 89–92. (enlace)
- Restall, R., C. Rodner & M. Lentino (2006). Birds of northern South America. An identification guide. Volume 1. Species accounts. Yale University Press, New Haven, Connecticut. 1536 pp. ISBN 9780300125047
- Ridgely, R. S. & P. J. Greenfield (2001) The Birds of Ecuador: status, distribution and taxonomy. Vol 1 y 2. Cornell University Press, Ithaca, NY. 880 pp. ISBN 9780801487200
- Ridgely, R. S. & J. A. Gwynne (1989). A Guide to the Birds of Panama with Costa Rica, Nicaragua, and Honduras. Second edition. Princeton University Press, Princeton, New Jersey, USA. 534 pp. ISBN 0-691-02512-6
- Ridgway, R. (1916). The birds of North and Middle America. Part VII. Bulletin of the United States National Museum 50, part 7. (enlace)
- Rios, M., G. Londoño & L. Biancucci (2008). Notes on birds that follow army ants in the northern Andes. Ornitología Neotropical, 19:137–142. (enlace)
- Roesler, I. & M. G. Agostini (2012). Inventario de los vertebrados de la Reserva Natural Punta Lara, Provincia de Buenos Aires, Argentina. (enlace)
- Ronchi-Virgolini, A. L., Lorenzón, R. E., Beltzer, A. H. & J. M. Alonso (2010). Ensambles de aves del Parque Nacional Pre-Delta (Entre Ríos, Argentina): análisis de la importancia ornitológica de distintas unidades ambientales. Hornero, 25 (1): 27-40. (enlace)
- Rowley, J. S. (1984). Breeding records of land birds in Oaxaca, Mexico. Proceedings of the Western Foundation of Vertebrate Zoology ,2 (3): 76–221. ISSN 0511-7550
- Russell, S. M. (1964). A distributional study of the birds of British Honduras. Ornithological Monographs number 1. American Ornithologists’ Union, Washington, D.C. (enlace)
- Salvador, S. A. & A. Bodrati (2013). Aves víctimas del parasitismo de moscas del género Philornis en Argentina. Nuestras aves, 58: 16-21. (enlace)
- Salvador, S. A. (2016). Distribución reproductiva de las aves de Argentina y sus territorios. Recopilación inédita. Córdoba, Argentina. (enlace)
- Salvador, S. A., Salvador, L. A., Ferrari, C. & S. Vitale (2016). Listado de aves de la provincia de Córdoba, Argentina. «Birds Checklist”. Córdoba, Argentina. (enlace)
- Salvador, S. A., L. A. Salvador & C. Ferrari (2017). Aves de la provincia de Córdoba. Distribución e Historia Natural. Ed. DP. Argentina. SA.
- Schubart, O., A. C. Aguirre & H. Sick (1965). Contribuição para o conhecimento da alimentação das aves brasileiras. Arquivos de Zoologia do Estado de São Paulo, 12: 95–249. (enlace)
- Schulenberg, T. S., D. F. Stotz, D. F. Lane, J. P. O’Neill, and T. A. Parker III. (2007). Birds of Peru. Princeton University Press, Princeton, New Jersey. 664 pp. ISBN 978-0-691-13023-1
- Selander, R. & D. R. Giller (1959). The avifauna of the Barranca de Orlatos, Jalisco, Mexico. Cóndor, 61: 210-222. (enlace)
- Short, L. L. (1975:231). A zoogeographic analysis of the South American Chaco avifauna. Bulletin of the American Museum of Natural History, 154 (3): 165–352. (enlace)
- Sick, H. (1985). Ornitologia Brasileira. Uma Introdução. Universidad de Brasília, Brasília, Brazil. 912 pp. ISBN 85-209-0816-0
- Sick, H. (1993). Birds in Brazil: A Natural History. Princeton University Press, Princeton, NJ, USA. (enlace)
- Skutch, A. F. (1966). Life history notes on three tropical American cuckoos. Wilson Bull., 78 (2): 139–165. (enlace)
- Skutch, A. F. (1983). Birds of Tropical America. University of Texas Press, Austin, TX, USA. (enlace)
- Slud, P. (1964). The birds of Costa Rica: distribution and ecology. Bulletin of the American Museum of Natural History, 128: 1–430. (enlace)
- Snyder, D. E. (1966). The Birds of Guyana. Peabody Museum, Salem, MA, USA. (enlace)
- Sorenson, M. D. & R. B. Payne (2005). A molecular genetic analysis of cuckoo phylogeny. In The Cuckoos (R. B. Payne, Editor). Oxford University Press, Oxford, UK. pp. 68–94
- Steullet, A. B. & E. A. Deautier (1935). Catálogo sistemático de las aves de la República Argentina. Imprenta y Casa Editora» Coni». Volumen 1, Nº 1 de Universidad Nacional de La Plata. Instituto del Museo. Obra del cincuentenario del Museo de La Plata, República Argentina.
- Stiles, F. G. & A. F. Skutch (1989). A Guide to the Birds of Costa Rica. Christopher Helm, London, UK. (enlace)
- Stotz, D. F., J. W. Fitzpatrick, T. A. Parker & D. K. Moskovits (1996). Neotropical Birds: Ecology and Conservation. University of Chicago Press, Chicago, IL, USA. ISBN 0-226-77630-1
- Terborgh, J. W., J. W. Fitzpatrick & L. Emmons (1984). Annotated checklist of bird and mammal species of Cocha Casu Biological Station, Manu National Park, Peru. Fieldiana Zoology, 21: 1–29. (enlace)
- Terborgh, J. W., Robinson, S. K., Parker, T. A., Munn, C. A. & N. Pierpont (1990). Structure and organization of an Amazonian forest bird community. Ecological Monographs, 60: 213–238. (enlace)
- Todd, W. E. C. & M. A. Carriker, Jr. (1922). The birds of the Santa Marta region of Colombia: a study in altitudinal distribution. Annals of the Carnegie Museum, 14. (enlace)
- Verea, C., A. Solórzano, M. Díaz, L. Parra, M. A. Araujo, F. Antón, O. Navas, O. J. L. Ruiz & A. Fernández-Badillo (2009). Registros de actividad reproductora y muda en algunas aves del norte de Venezuela. Ornitología Neotropical, 20 (2):181–201. (enlace)
- Vides Almonacid, R. (1993). Estudio comparativo de las taxocenosis de aves de los bosques montanos de Sierra de San Javier, Tucumán, Argentina. Bases para su manejo y conservación- Tesis doctoral, Facultad de Ca. Naturales, Universidad Nacional de Tucumán, Argentina; 347 pp.
- Waldrigues, A., Ferrari, I.& A. F. Neto (1983). Estudio cariotípico em duas espécies de cuculiformes americanos (Aves). [Caryotypic studies of two species of American Cuculiformes (Aves)]. Acta Amazonica, 13 (1): 37–50. (enlace).
- Wetmore, A. (1939). Observations on the birds of northern Venezuela. Proceedings of the United States National Museum 87:173–260. (enlace)
- Wetmore, A. (1968). The Birds of the Republic of Panama. Part 2. Columbidae (Pigeons) to Picidae (Woodpeckers). Smithsonian Miscellaneous Collections 150. Smithsonian Institution, Washington, DC, USA. (enlace)
- White, E. W. (1882). Notes on birds collected in the Argentine Republic. Proc. Zool. Soc. London: 591-629. (enlace)
- Willis, E. O. & E. Eisenmann (1979). A revised list of birds of Barro Colorado Island, Panama. Smithsonian Contributions to Zoology, 291: 1–31. (enlace)
- Willis, E. O. (1983). Piaya cuckoos (Aves, Cuculidae) as army ant followers. Revista Brasileira de Zoologia, 43: 29-32.
- Yzurieta, D. (1995) Manual de reconocimiento y evaluación ecológica de las aves de Córdoba. Argüello, Argentina: Ministerio de Agricultura, Ganadería y Rec. Renovables. 438 pp.
- Zapata, A. R. P. (1975). Aves observadas en la proximidad de la confluencia de los ríos Uruguay y Gualeguaychú, provincia de Entre Ríos. Hornero, 11 (4): 291-304 (enlace)
- Zapata, A. R. P. (1977). Aves observadas en la proximidad de la confluencia de los ríos Uruguay y Gualeguaychú, provincia de Entre Ríos. Hornero, 11 (5): 387-403. (enlace)
- Zimmer, J. T. (1930). Birds of the Marshall Field Peruvian Expedition, 1922–1923. Field Museum of Natural History Zoological Series, 17: 233–480. (enlace)
-
-
Tingazú | Conservación
ESTADO DE CONSERVACIÓN SEGÚN BIRDLIFE INTERNACIONAL: PREOCUPACIÓN MENOR.Esta especie tiene un rango extremadamente grande, y por lo tanto no se acerca a los umbrales de Vulnerable bajo el criterio de tamaño del rango (Extensión de Ocurrencia <20,000 km2 combinado con un tamaño del rango decreciente o fluctuante, extensión/calidad del hábitat, o tamaño de la población y un pequeño número de ubicaciones o fragmentación severa). A pesar del hecho de que la tendencia de la población parece estar disminuyendo, no se cree que la disminución sea lo suficientemente rápida como para acercarse a los umbrales de Vulnerabilidad bajo el criterio de tendencia de la población (>30% de disminución durante diez años o tres generaciones). El tamaño de la población no ha sido cuantificado, pero no se cree que se acerque a los umbrales de Vulnerable bajo el criterio de tamaño de la población (<10,000 individuos maduros con una disminución continua estimada en >10% en diez años o tres generaciones, o con una estructura de población específica). Por estas razones la especie es evaluada como Preocupación Menor.
La población mundial no ha sido cuantificada debido a recientes divisiones taxonómicas (Birdlife International, 2025).La abundancia relativa del cuco ardilla se clasifica como «poco común a bastante común» en Ecuador (Ridgely & Greenfield, 2001), «bastante común» en Trinidad (ffrench, 1991:190), «común a bastante común» en México (Howell & Webb, 1995:348) o «común» en Honduras (Monroe, 1968:330), Costa Rica (Stiles & Skutch, 1989:186), Panamá (Ridgely & Gwynne, 1989:183), Venezuela (Hilty, 2003:352), Colombia (Hilty & Brown, 1986:221) y Surinam (Haverschmidt & Mees, 1967:151).
Estado de conservación en Argentina (SIB)
AvA-SADS-NA: No amenazada según AvA SAyDS 2008 (Aves Argentinas y Secretaría de Ambiente y Desarrollo Sustentable). López-Lanús, B., P. Grilli, E. Coconier, A. Di Giacomo y R. Banchs. Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas /AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina 1 ed. AOP: Buenos Aires, Argentina
Res 348 SA-NA: No Amenazada según SAyDS Res 348 (Secretaría de Ambiente y Desarrollo Sustentable). Categorización de aves autóctonas de Argentina. Lista de aves no registradas, endémicas y exóticas de Argentina 2010.Efectos de la actividad humana
La actividad humana tiene poco efecto directo sobre el cuco ardilla, salvo los efectos locales de la destrucción del hábitat. Sin embargo, toleran relativamente bien las perturbaciones del hábitat, siempre que se mantenga vegetación leñosa de altura moderada (Fitzgerald et al., 2020). -
Tingazú | Distribución
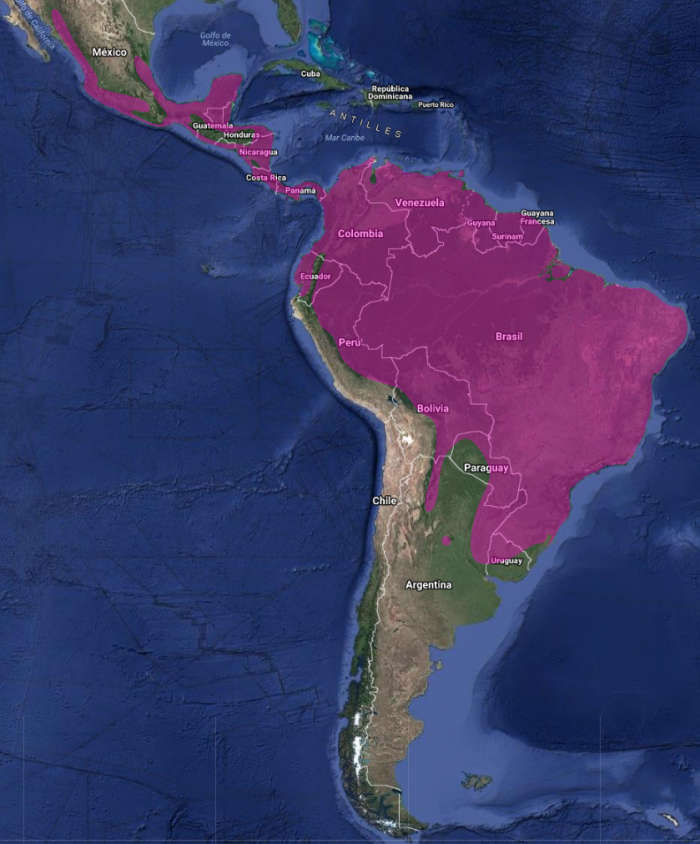
InfoNatura: Animals and Ecosystems of Latin America 2007 Especie endémica de América presente en la mayor parte de la América tropical continental, desde el norte y centro de México hasta el sur de Panamá y en América del Sur desde el suroeste de los Andes hasta el noroeste de Perú y, al este de los Andes, en Trinidad y desde el norte hasta el sur de Argentina (Skutch, 1966:139). En América Central habita tanto distritos húmedos como secos, desde el nivel del mar hasta zonas montañosas.
Su distribución altitudinal es notablemente amplia, ocupando desde tierras bajas hasta zonas montañosas. Sin embargo, generalmente es más abundante en altitudes bajas y raro por encima de los 1200-1400 msnm (Schafer & Phelps, 1954:61). Es una especie residente en toda su área de distribución (Howell & Webb, 1995:348).
En México alcanza los 2000 msnm (Friedmann et al., 1950:133; Howell & Webb, 1995:348; Skutch, 1966:139); en Guatemala y Costa Rica, los 2450 msnm (Land, 1962:272), y hasta los 2000 msnm en las laderas húmedas del volcán Irazú. En otras zonas como Vara Blanca, no parece ser residente por encima de los 1600 msnm (Land, 1962:272; Stiles & Skutch, 1989:186); en Panamá, más de 1800 msnm (Ridgely & Gwynne, 1989:183); en Colombia, hasta aproximadamente 2000 msnm, y en menor número hasta los 2700 msnm, incluyendo la península de La Guajira y el sur de Florida (Hilty & Brown, 1986:221). En Venezuela, se observa hasta los 2500 msnm (Schafer & Phelps, 1954:61), aunque generalmente por debajo de los 1200 msnm al norte del Orinoco y hasta los 1800 msnm al sur de este río (Hilty, 2003:352). En Ecuador, se registra regularmente hasta los 1500 msnm, en menor cantidad hasta los 2000–2200 m, y ocasionalmente hasta los 2500 m e incluso 2700 m en Loja (Ridgely & Greenfield, 2001). En Perú, habita hasta los 2800 msnm, siendo la especie del género Piaya más común y ampliamente distribuida en bosques húmedos y semihúmedos del oriente peruano (Schulenberg et al., 2007:184). En El Salvador, también ha sido documentada (Dickey & van Rossem, 1938:213–215).Hábitat
El Tingazú es una especie que ocupa diversos hábitats boscosos dentro de su rango de distribución. Se encuentra en bosques húmedos y semihúmedos, bosques caducifolios, bordes de bosques, bosques secundarios y plantaciones, así como en áreas con árboles dispersos en campos abiertos (Haverschmidt, 1967:151; Hilty & Brown, 1986:220; Binford, 1989:127; Stiles & Skutch, 1989:186; Howell & Webb, 1995:348; Ridgely & Greenfield, 2001). También está presente como residente permanente en las regiones Atlántica y Pacífica, en bosques siempreverdes tropicales, semideciduos, deciduos y pantanosos del Pacífico, así como en bosques de galería dentro de zonas de matorral tropical árido, como en la cuenca del río Verde (San Pedro Juchatengo) y en áreas de matorral subtropical árido cerca de Huajuapan de León, desde el nivel del mar hasta los 1600 msnm (Binford, 1989:127).
Evita los bosques densos e ininterrumpidos, mostrando predilección por áreas abiertas o semiabiertas con árboles dispersos. Se lo observa frecuentemente en zonas cultivadas, setos, cafetales con sombra, pastizales, patios con árboles espaciados y claros abandonados con vegetación densa. Se alimenta en los bordes del bosque, entre enredaderas o en las copas de los árboles próximos, aunque evita la vegetación sombría y cerrada (Skutch, 1966:139). En regiones semidesérticas, se lo observa principalmente en árboles altos ubicados cerca de cursos de agua o en zonas bajas. En Trinidad es considerado residente común, distribuido tanto en bosques como en áreas cultivadas semiabiertas (ffrench, 1991:190).
En Argentina, específicamente en la Reserva El Bagual, el cuco ardilla habita isletas de urundayzal y bosques ribereños, ocupando principalmente los estratos alto y medio de la vegetación. Su abundancia relativa en esta región se considera poco común (Di Giácomo, 2005:290).En Argentina, el Tingazú se distribuye en dos subespecies principales. Piaya cayana macroura Gambell, 1849 se encuentra en las provincias de Misiones, Corrientes, Entre Ríos, este de Formosa y Chaco, norte y este de Santa Fe, este de Córdoba y noreste de Buenos Aires. Por su parte, Piaya cayana mogenseni J. L. Peters, 1926 habita en Tucumán, Santiago del Estero, Catamarca, Salta y Jujuy (de la Peña, 2016:328; de la Peña, 2020:89; Antelo & Brandán Fernández, 2013:91). (Citas y observaciones en los enlaces por provincias)
-
Tingazú | Comportamiento
Según Fitzgerald et al. (2020), buscan alimento principalmente en el estrato medio y el dosel del bosque, donde, pese a su tamaño considerable, pueden pasar desapercibidos. Se desplazan saltando entre ramas y su movimiento, junto con su coloración, recuerda al de una ardilla, lo que explica su nombre común en inglés. Frecuentemente se trasladan de un árbol a otro mediante planeos prolongados interrumpidos ocasionalmente por breves aleteos.
Estos cucos pueden alimentarse ocasionalmente en asociación con hormigas guerreras. Willis & Eisenmann (1979:14) indicaron que P. minuta, P. melanogaster y P. cayana rara vez siguen a estas hormigas para capturar presas espantadas, ya que están mejor adaptadas al forrajeo entre el follaje que cerca del suelo. Willis (1983:29) añadió que la mayoría de los registros de esta conducta corresponden a P. cayana, especialmente en bordes de bosque o en el interior de bosques panameños al final de la temporada lluviosa y reproductiva. En la isla Barro Colorado, Panamá, esta conducta es estacional, con mayor frecuencia entre noviembre y diciembre (Willis, 1983:29). Las observaciones se concentran en individuos buscando alimento sobre enredaderas de lianas o en el dosel de árboles secundarios, más que en el suelo o sotobosque.
En cuanto a las interacciones reproductivas, la cópula es precedida por alimentación nupcial, sin exhibiciones adicionales ni vocalizaciones (Skutch, 1966:142). En Costa Rica, Skutch (1966) observó adultos alimentando a sus parejas, conducta que en ocasiones antecedía inmediatamente a la cópula, durante la cual ambos sostenían insectos en sus picos.
Respecto a la estructura social, son generalmente solitarios, aunque habitualmente se observan en parejas y ocasionalmente forman pequeños grupos (ffrench (1991:191; Fitzgerald et al., 2020). Pueden seguir bandadas mixtas, aunque a menudo se alimentan de manera independiente. Skutch (1966:140) destacó que no realizan vuelos prolongados, prefiriendo desplazarse entre árboles mediante saltos o cortos planeos (de la Peña, 2016:327). Para ganar altura, ascienden gradualmente de rama en rama, pero para descender ejecutan largos planeos con las alas extendidas y la cola desplegada, emitiendo sonidos metálicos. También señaló que no forman bandadas, sino que viven en parejas todo el año, aunque los individuos no permanecen juntos constantemente; suelen separarse por varios árboles y mantener contacto mediante vocalizaciones.
VOCALIZACIONESEl cuco ardilla (Piaya cayana) vocaliza con poca frecuencia, pero presenta un repertorio amplio y distintivo (Fitzgerald et al., 2020). Sus llamadas incluyen un reclamo áspero descrito como “jicaro”, “eee-kah” o “kweep-kweep” (Skutch, 1966:141), “kikerah” o “geep-kareer” (Ridgely & Gwynne, 1989:183), y “chek-e-rehr” (Howell & Webb, 1995:348). También emite un llamado bisílabo “eee-kah” o “chick-kwah” (Skutch, 1966:141; Howell & Webb, 1995:348), y un canto explosivo “KEEK! wahh” (Lane in Schulenberg et al., 2007:184).
Durante el vuelo produce notas cortas como “trrt-trrt-trrt” (Ridgely & Gwynne, 1989:183) o “djet!-djet!” (Hilty, 2003:351), y un cascabeleo seco y gutural (Howell & Webb, 1995:348). Su canto consiste en una serie de notas repetidas “whip-whip-whip…” o “wheep” que pueden ser fuertes o suaves (Skutch, 1966:141; Willis & Eisenmann, 1979:14).
Belton (1984:542) lo describió como una voz alta y abrupta, que incluye gruñidos ásperos y sonidos nasales. de la Peña, (2016:327) y Straneck (in litt.) mencionan secuencias de silbidos cortos “juit-juit-juit” y un llamado fuerte “chrrrrr”. Estas vocalizaciones cumplen funciones tanto territoriales como de comunicación entre los individuos.ALIMENTACIÓNSegún Fitzgerald et al. (2020), el tingazú o cuco ardilla (Piaya cayana) es principalmente insectívoro. Aunque los datos disponibles son escasos, se han registrado como presas diversas orugas —incluidas especies con pelos urticantes—, insectos palo, saltamontes, escarabajos, avispas, abejas, hormigas grandes, cigarras, hemípteros, odonatos, insectos no identificados y arañas de gran tamaño (Todd & Carriker, 1922:212; Dickey & van Rossem, 1938:215; Schubart et al., 1965:151). También se ha documentado el consumo de restos quitinosos de escarabajos y pulpa de frutos pequeños (Blake & Hanson, 1942:529).
Adicionalmente, esta especie puede capturar pequeños vertebrados. Wetmore (1968:118) registró la depredación ocasional de lagartijas pequeñas, y Cintra & Sanaiotti (1990:470) observaron a un individuo capturando y matando una rana arbórea (Osteocephalus taurinus), la cual no fue consumida, probablemente debido a su tamaño (10 cm), lo que sugiere que podrían alimentarse de ranas más pequeñas. También se ha planteado la posible depredación de huevos de aves, aunque no existen observaciones directas que lo confirmen (Gibbs, 1991). En cuanto al consumo de frutos, se han registrado especies como Plinia trunciflora (guapurú – ibá porú – îwa purú – yvapuru) (Bodrati, 2006), Allophylus edulis (chalchal, cocó, cocú, kokú, wakú) (Bodrati & Haene, 2008) y semillas de Cassia chrysocarpa (Schubart et al., 1965:151).
Skutch (1966:140) afirmó que el cuco ardilla es, en su experiencia, exclusivamente insectívoro, y describió su método de caza como una combinación de movimientos pausados y escrutinio minucioso, seguidos de saltos repentinos, a veces de gran longitud. El ave examina lentamente el follaje con sus ojos rojos, y una vez que detecta una presa, la captura con rapidez. Posteriormente, la golpea contra una rama o la tritura con el pico antes de ingerirla. Este comportamiento fue reiterado por Skutch (1983:55–56), quien también destacó la captura de insectos palo de hasta 10–12 cm de longitud.
Todd & Carriker (1922:212) señalaron que la dieta de Piaya columbiana, especie en ocasiones considerada conespecífica con P. cayana, se compone casi exclusivamente de insectos palo (Phasmatodea), particularmente los de mayor tamaño. No obstante, en Centroamérica, P. cayana consume una mayor variedad de insectos, incluidas orugas verdes y polillas, a pesar de que algunas orugas pueden poseer espinas venenosas irritantes.
Registros adicionales confirman la ingestión de diversas órdenes de insectos, como Coleoptera (Chrysomelidae), Hemiptera (Pentatomidae), Homoptera (Cicadidae, Fulgoridae), Hymenoptera (Pergidae, Meliponidae), Lepidoptera, Orthoptera (Acridiidae, Tettigoniidae), así como arácnidos (Araneidae) (Repenning et al., 2009; Schubart et al., 1965:151; Haverschmidt & Mees, 1967:151; Wetmore, 1968:118; de la Peña, 2016:327).REPRODUCCIÓNLa biología reproductiva ha sido ampliamente estudiada en diversas regiones de su distribución (Payne, 2005:334; Fitzgerald et al., 2011).
El cuco ardilla construye nidos en árboles, arbustos o vegetación densa y baja (Skutch, 1966:143; Rowley, 1984). El nido consiste en una plataforma suelta de ramas gruesas cubierta por hojas, algunas frescas, y mide aproximadamente 15–18 cm de diámetro y 6–9 cm de profundidad (Skutch, 1966:143; 1983:57). La construcción es realizada por ambos miembros de la pareja, uno aportando materiales y el otro acomodándolos (Skutch, 1966:144).
La puesta suele ser de dos huevos blancos, ocasionalmente manchados por el material del nido. Las dimensiones registradas son 34,6 ± 1,04 × 25,3 ± 0,88 mm (n=8) (Cherrie, 1892:326; Skutch, 1966:146; Rowley, 1984). En algunos casos se han reportado hasta tres huevos y tonalidades grisáceas recubiertas de material calcáreo (de la Peña, 2016:327). La incubación, compartida por ambos sexos, dura 18–19 días (Skutch, 1966:149).
Ambos progenitores alimentan a los polluelos con pocas presas grandes por la mañana, mantienen el nido limpio y aportan hojas frescas (Skutch, 1966:149; 1983:58). Los juveniles comienzan a salir gradualmente del nido hacia los ocho días, permaneciendo cerca en ramas próximas antes de independizarse progresivamente (Skutch, 1966:149; 1983:58).
El cortejo incluye alimentación entre la pareja, en ocasiones seguida de cópula (Skutch, 1966:142). Este comportamiento ha sido observado en Costa Rica y se caracteriza por el intercambio de insectos durante la monta. La reproducción ocurre en distintas épocas según la región: de enero a octubre en Trinidad (ffrench, 1991:191), y entre noviembre y enero en Argentina (de la Peña, 2016:327).
Los nidos han sido registrados en diversos hábitats y alturas: desde 0,7 hasta 12 m sobre el suelo, en árboles, helechos, bambúes, palmeras y ombúes (Skutch, 1966:143; de la Peña, 2016:327; Di Giacomo, 2005:289-290). En Argentina se documentaron nidos en Salta, Formosa y Misiones, con estructuras semiesféricas de ramitas y hojas (de la Peña, 2016:327).
Depredadores incluyen Accipiter bicolor y Bubo virginianus (Di Giacomo, 2005:289). Además, se han registrado casos de parasitismo por moscas del género Philornis (Bodrati en Salvador & Bodrati, 2013). -
Tingazú | Descripción

© Sebastián Marjanov. Tingazú (P. c. macroura). Chacra del Agua. El Soberbio. 4 de Febrero 2022. Algunos derechos reservados 
© Alejandro Bayer Tamayo. Piaya cayana (Cuco ardilla). Armenia, Colombia. Febrero de 2017. Algunos derechos reservados 
© Barloventomágico. Piscua. (Piaya cayana mehleri) Sabaneta, El Hatillo, Estado Miranda, Norte de Venezuela. Algunos derechos reservados. De gran tamaño, y hábitos arborícolas, caracterizadas por una cola muy larga y escalonada. Presentan partes superiores de color castaño rojizo, garganta y pecho beige, y vientre gris claro. La cola es rojiza por encima, mientras que la superficie inferior de las timoneras muestra tonalidades negruzcas con amplias puntas blancas. El nombre común “cuco ardilla” deriva tanto de su coloración como de sus movimientos entre la vegetación, los cuales recuerdan a los de una ardilla (de la Peña, 2016: 326; Fitzgerald et al., 2020)
La descripción morfológica se basa principalmente en Ridgway (1916:45), Dickey & van Rossem (1938:213-215) y Wetmore (1968:116), con especial referencia a Piaya cayana thermophila. Los adultos presentan partes superiores de castaño a castaño rojizo, generalmente más pálidas en la cabeza. Las remeras poseen puntas de color marrón grisáceo. La cola, larga y graduada, es rojiza dorsalmente, con una banda subterminal negra y una mancha terminal blanca en cada timonera; ventralmente, las timoneras son de color negro parduzco con puntas blancas. La garganta y el pecho son canela; el abdomen es gris, con variaciones hacia tonos pizarra o negro pizarroso en los flancos, el crissum y las cubiertas subcaudales. El iris es rojo carmín o rojo oscuro (Ridgway, 1916:45; Haverschmidt, 1967:151; Wetmore, 1968:116). El pico es amarillo verdoso (Haverschmidt, 1968:151) y los tarsos y dedos muestran color gris azulado o gris plomo (Ridgway, 1916:45; Haverschmidt, 1968:151).
El plumaje definitivo adulto se adquiere mediante una muda prebásica completa. En El Salvador, esta muda se observa principalmente entre julio y septiembre (Dickey & van Rossem, 1938:213).
El color de la piel orbital desnuda presenta variación geográfica. Desde el sur de México hasta el noroeste de Sudamérica, así como en el oeste de los Andes, el anillo ocular es verde amarillento a verde oliva claro (subespecies mexicana, thermophila y nigricrissa (Ridgway, 1916:45; Wetmore, 1968:116; Howell & Webb, 1995:348; Ridgely & Greenfield, 2001; Schulenberg et al., 2007:184). En el norte de Colombia, noroeste y norte de Venezuela (subespecies mehleri, circe), y en Trinidad (insulana), también se ha registrado coloración amarilla (Restall et al., 2006; ffrench, 1991:190). Desde Venezuela al sur del río Orinoco y hacia el extremo meridional de la distribución, el anillo ocular es carmesí (Haverschmidt, 1967:151; Wetmore, 1968:116; Belton, 1984:541; Hilty & Brown, 1986:220; Ridgely & Greenfield, 2001; Hilty, 2003:351; Schulenberg et al., 2007:184).En cuanto a las medidas, se basan en las descripciones de P. c. thermophila
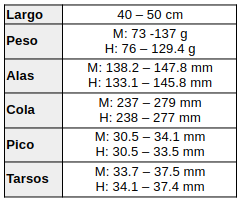
(Wetmore, 1968:116; Ridgway, 1916:45; Howell & Webb, 1995:348; Hilty & Brown, 1986:220; Hilty, 2003:351; Payne, 2005:332) Para la subespecie nominal cayana, tanto machos como hembras registran en Surinam una media de 95 g (machos: 84–103 g, n=10; hembras: 82–103 g, n=6) (Haverschmidt & Mees, 1967:151).
-
Tingazú | Taxonomía
Se han descrito diversas subespecies de Piaya cayana, reconociéndose actualmente catorce (Payne, 2005:332). La delimitación de estas subespecies se fundamenta principalmente en diferencias de tamaño corporal y coloración del plumaje, aunque no siempre es evidente la correspondencia entre estas variaciones y el color del anillo ocular. En este sentido, se considera necesaria una revisión moderna y exhaustiva de la variación geográfica de Piaya cayana, basada en múltiples caracteres, incluida la evidencia genética.
La mayoría de las subespecies presumiblemente se intergradan en las zonas donde sus áreas de distribución entran en contacto. No obstante, en el sur de México, Binford (1989:128) observó especímenes intermedios entre P. c. thermophila y P. c. mexicana, pero destacó que lo abrupto y raro de dicha intergradación sugieren que estas dos formas podrían representar especies distintas.
Con base en un análisis filogenético de secuencias de ADN mitocondrial, Sorenson & Payne (2005:91) propusieron la división de la familia en cinco subfamilias. Piaya fue ubicada dentro de Cuculinae, subfamilia de distribución global, y su género hermano identificado como Coccyzus, de distribución neotropical. Tradicionalmente, Piaya incluía tres especies; sin embargo, los autores demostraron que Coccycua minuta forma un clado junto con dos especies anteriormente asignadas a Coccyzus. Estas tres especies constituyen actualmente el género Coccycua. En consecuencia, Piaya comprende solo dos especies válidas: P. cayana y P. melanogaster (Alma de gato negro).
El siguiente resumen de la distribución y los caracteres de las subespecies se basa en Peters (1940:44) y Payne (2005:332):- Piaya cayana mexicana, Swainson 1827; (monotípica)
Se encuentra en la vertiente del Pacífico de México, al sur del Istmo de Tehuantepec.
Color castaño rojizo más claro en el haz; envés de las timoneras rojizo, con barras subterminales negras y puntas blancas anchas. - Piaya cayana thermophila, Sclater 1859; (monotípica)
Este de México, a través de Centroamérica hasta Panamá y el noroeste de Colombia.
Color castaño rojizo relativamente oscuro en el dorso; vientre y cubiertas subcaudales de gris oscuro a negro; parte inferior de las timoneras negra; puntas blancas de las timoneras relativamente estrechas. - Piaya cayana nigricrissa, Cabanis 1862; (monotípica)
Oeste de Colombia (y al este hasta las laderas de los Andes centrales), oeste de Ecuador y noroeste de Perú.
Similar a la thermophila, pero con plumaje más oscuro; vientre y cubiertas subcaudales negruzcas. - Piaya cayana mehleri, Bonaparte 1850
Se encuentra en el noreste de Colombia, desde el Golfo de Urabá hasta el valle del Magdalena y la vertiente occidental de los Andes orientales, al este a lo largo de la costa norte de Venezuela hasta la Península de Paria. - Piaya cayana mesura, Cabanis & Heine 1863
Colombia al este de los Andes, este del Ecuador y noreste del Perú. - Piaya cayana circe, Bonaparte 1850
Se encuentra en Venezuela, al sur del lago Maracaibo. - Piaya cayana cayana, Linné 1766
Este y sur de Venezuela a través de las Guayanas y ribera sur a norte del bajo Amazonas. - Piaya cayana insulana, Hellmayr 1906
En Trinidad. - Piaya cayana obscura, Snethlage 1908
Centro de Brasil, sur del Amazonas hasta el alto Ji-Paraná, este de Perú y norte de Bolivia. - Piaya cayana hellmayri, Pinto 1937
En Brasil al sur del Amazonas, desde Santarem al este hasta la desembocadura del Amazonas, y costa norte de Maranhão. - Piaya cayana pallescens, Cabanis & Heine 1863
Este de Brasil. - Piaya cayana cabanisi, Allen 1893
Sur y centro de Brasil (Centro de Mato Grosso y Goiás a norte de São Paulo). - Piaya cayana macroura, Gambel 1849
Sureste de Brasil a Paraguay, noreste de Argentina y Uruguay.
Parte superior oscura; vientre y cubiertas subcaudales negruzcas. - Piaya cayana mogenseni, Peters 1926
Sur de Bolivia a noroeste de Argentina.
Parte superior rojiza más clara; garganta rojiza; parte inferior gris oscuro (no negruzca).
- Piaya cayana mexicana, Swainson 1827; (monotípica)
-
Tingazú (Piaya cayana)

Planches enluminées d’histoire naturelle. Martinet, François Nicolas, Buffon, Georges Louis Leclerc, Daubenton, Edme-Louis. (1765-1783?); Vol III: Lámina 211 El Tingazú (Piaya cayana) es una especie común que ocupa una amplia gama de hábitats boscosos, incluyendo ambientes naturales y modificados, a lo largo de un extenso gradiente de humedad. Se le observa frecuentemente en actividad arbórea, desplazándose en busca de artrópodos. Presenta un alto grado de variación morfológica, con catorce subespecies diferenciadas principalmente por la coloración de distintas partes del cuerpo. Algunas de estas subespecies podrían constituir especies independientes, dado su escaso grado de integración con subespecies adyacentes.
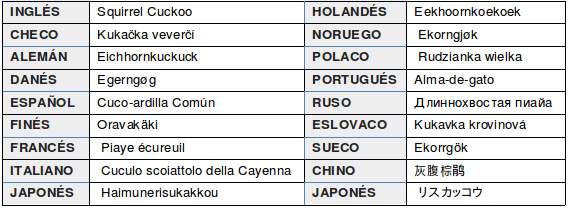
Nombres comunes: Cuco ardilla (Colombia, Costa Rica, Perú, Nicaragua, México); Alma de Gato (Brasil); Tingazú (Argentina, Bolivia y Paraguay); Pirincho del Monte y Urraca Canela (Uruguay); Piscua (Venezuela); Pájaro León (Honduras); «Mbaymy-Tytyrú, guaimí-tiriríh, güira-payé, guaymingué, tingasú, o guaimí-tyryrú» en guaraní; «Cachó« en guayaquí; Anó del monte, Gallo del monte, Gato del monte, Güira-payé, Pirincho marrón, Anó colorado, Urraca colorada, Urraca real, Cola de chala; Alma de caboclo, o tabo de paia (Brasil) (de la Peña, 2016:326; Chébez, 1996:132).
En otros idiomas
Breves apuntes históricos
- Historia Naturalis Brasiliae. Willem Piso, George Marcgrave (1648): 216 (Lámina) (Atingaçu camacu)
- Méthode contenant la division des oiseaux en ordres, sections, genres, especes & leurs variétés. Brisson, Mathurin-Jacques, Martinet, François Nicolas. Parisiis: Ad Ripam Augustinorum, apud Cl. Joannem-Baptistam Bauche, bibliopolam, ad Insigne S. Genovesae, & S. Joannis in Deserto, 1760. Vol IV: lámina VIII (Fig.2) (Coucou de Cayenne)
- Planches enluminées d’histoire naturelle. Martinet, François Nicolas, Buffon, Georges Louis Leclerc, Daubenton, Edme-Louis. Paris?:s.n., 1765-1783? Vol III: Lámina 211 (Coucou de Cayenne)
- Peces y aves del Paraguay Natural (1767). José Sánchez Labrador. Pp: 354; Lámina (Ave tingazo)
- Apuntamientos para la Historia Natural de los Páxaros del Paragüay y Río de la Plata (1805). Félix de Azara. Vol II: 265 (Tingaru)
- Dictionnaire d’histoire naturelle. Paris, Chez Deterville, 1816-19. Vol VIII: 275-277 (Le coulicou piaye – Coccyzus macrocercus)
- Encyclopédie méthodique. Bonnaterre, abbé (Pierre Joseph), Bénard, Robert, Vieillot, L. P. (Louis Pierre). A Paris: Chez Mme. veuve Agasse …,1823. Vol III: 1347 (Le coulicou piaye – Coccyzus macrocercus)
- A general history of birds. Latham, John. Winchester [England]1821-1828; Vol III: 322-323 (Cayenne cuckow)
- Tableau méthodique des ordres, sous-ordres, familles, tribus, genres, sous-genres et races d’oiseaux. Lesson, R. P. (René Primevère), Prêtre, J. G. Bruxelles: Chez F.G. Levrault; 1831.Vol I (1831): 140 (Piaya cayana)
- Argentine ornithology. A descriptive catalogue of the birds of the Argentine Republic. Sclater, Philip Lutley, Hudson, W. H. London: R. H. Porter, 1888-89. Vol II: 36-37 (Piaya cayana)
- Catalogue of the birds in the British Museum. British Museum (Natural History). Department of Zoology. [Birds]. Vol XIX (1891): 373-377 (Piaya cayana)
- De vogels van Guyana (Suriname, Cayenne en Demerara). Penard, Frederik Paul. Paramaribo: E.P. Penard; [1908-10]; Vol II: 26