Es un ave de hábitos solitarios, comportamiento pasivo y actividad predominantemente crepuscular. Se desplaza principalmente en los estratos medios e inferiores de la selva paranaense, donde suele permanecer posada en posición vertical, balanceando lateralmente su larga cola péndula. La defensa del territorio se realiza mediante vocalizaciones graves y profundas. Por lo general solo o en parejas y se reproduce en taludes o bancos cercanos al nivel del suelo. Su vuelo es lento y ondulado, característica atribuida a la corta longitud de sus alas (Snow & Kirwan, 2020).
VOCALIZACIONES
La especie presenta una actividad vocal predominantemente crepuscular y nocturna, con mayor frecuencia antes del amanecer y al atardecer.
Sus vocalizaciones consisten en emisiones breves, guturales y de carácter profundo, transcripto fonéticamente como “bu.ru.bu.bup” o “jurrrr”, repetido de forma esporádica y con clara preferencia por las horas del crepúsculo (de la Peña, 2015:370; de la Peña, 2020:24; Straneck en de la Peña, 2016). Otros autores coinciden describiéndolas como burbujeos sonoros y graves, tales como “bubububububub” (Belton, 1984:583), o como llamados secos y ventrílocuos, incluyendo variantes como “búuuu”, “du-dúg-dug-dug-dug-dug” y “rrrrü” (Sick, 1997:477). Estas emisiones suelen ser más audibles en ambientes de selva densa y oscura, donde la especie habita preferentemente (Snow & Kirwan, 2020; de la Peña, 2020:24).
ALIMENTACIÓN
La dieta de la especie se compone principalmente de insectos grandes y sus larvas, arañas, moluscos terrestres, pequeños reptiles, mamíferos y aves, con un aporte ocasional de fruta. En el caso de los jóvenes en el nido, su alimentación incluye artrópodos (41,8%), frutas (32,8%), gusanos (7,4%), pequeños vertebrados como ranas y lagartijas (6,6%), moluscos (0,8%) y elementos no identificados (10,6%). Dentro de los artrópodos, predominan los insectos (74,5%), seguidos por diplópodos, siendo los más comunes los Coleóptera (10%), Orthoptera, como los insectos palo (30%), Hymenoptera, especialmente hormigas y, en menor medida, abejas (60%), y orugas (30%). A medida que los jóvenes se desarrollan, la proporción de artrópodos disminuye, mientras que el consumo de frutos, especialmente de Virola gardineri y una especie de Myrtaceae, aumenta significativamente en la fase de volantón (Alves et al., 1999:33). Además, ocasionalmente sigue a las hormigas arrieras, aprovechando presas perturbadas por ellas, y a veces se une a bandadas mixtas de forrajeo (Aleixo, 1997, en Snow & Kirwan, 2020).
REPRODUCCIÓN
Se caracteriza por anidar en madrigueras excavadas entre enero y abril, período en el que el suelo se mantiene húmedo debido a las lluvias estivales. La puesta ocurre probablemente entre septiembre y octubre, con registros de polluelos alimentados a comienzos de octubre y durante la segunda mitad de noviembre, así como de juveniles recién volantones a principios de diciembre (Alves et al., 1999:33). Asimismo, se ha documentado un nido activo, aunque sin huevos, en noviembre en el noreste de Argentina (Protomastro, 2001:19).
El nido consiste en una madriguera excavada en bancos de tierra o reutilizada a partir de cavidades abandonadas por otros animales, como armadillos (Tatú), cuya estructura puede diferir notablemente. Generalmente presenta un túnel de más de 1 m de longitud que culmina en una cámara sin revestimiento de aproximadamente 35 × 14 cm. En el Parque Nacional Iguazú (departamento Iguazú, Misiones), se describió un nido en el suelo con dos entradas de 10 a 12 cm de diámetro, una asociada a una antigua cueva de tatú; el túnel medía 2,8 m de longitud y la cámara terminal, ubicada a 25 cm de profundidad, presentaba dimensiones similares (de la Peña, 2016:24). Protomastro (2001:19) también describió un nido excavado en el techo de una gran cavidad posiblemente perteneciente a un tatú, con galerías inactivas cercanas correspondientes a ciclos reproductivos previos; la galería activa alcanzaba 2,8 m y poseía una segunda entrada situada a 0,8 m de la cámara, aparentemente utilizada exclusivamente para la atención del nido.
La puesta consta de dos a tres huevos y ambos sexos participan en la alimentación de las crías, con visitas al nido que oscilan entre 0 y 10 por hora; sin embargo, se ha observado que uno de los miembros de la pareja puede invertir más en el cuidado parental, mientras el otro compensa mediante una mayor dedicación a la defensa territorial y a comportamientos antidepredadores (Alves et al., 1999:33). Previo a ingresar a la madriguera, ambos adultos muestran un comportamiento cauteloso, posándose repetidamente en ramas cercanas e inspeccionando el entorno; durante este proceso, realizan movimientos pendulares de la cola, característicos de los momotos y particularmente intensos durante la época reproductiva (Alves et al., 1999:32–33).
A pesar de que se conoce que ambos miembros de la pareja reproductora excavan el túnel y nidifican en una cámara ubicada a lo largo de este, la información disponible sobre la ecología reproductiva de la especie sigue siendo limitada (Sick, 1997). Un diagrama detallado de la estructura del nido se presenta en Alves et al. (1999:32).
Según los siguientes autores: Belton, 1984:583; Sick, 1997:477; Snow, 2001:264;Snow & Kirwan, 2020; de la Peña, 2015:370; de la Peña, 2020:24.
Sexos similares.
Ambos sexos tienen la frente, corona y nuca castaño rojizas. Lores, plumas por encima del ojo (superciliares), lados de la cara y auriculares negras bordeados de turquesa.
Mejillas, garganta y parte superior del pecho color oliva-azulado.
Partes superiores verdes con reflejos amarillentos, volviéndose verde-oliva en espalda y lomo, incluidas las cubiertas alares y la cola. Cubiertas alares primarias y remeras verdes más azuladas-brillantes en las puntas.
Garganta y pecho verde oliva. En zona superior y central del pecho mechón de plumas elongadas de color negro con puntas verdes (a veces ausentes durante la muda). Banda en parte inferior del pecho rojo-canela y parte inferior del vientre y subcaudales verde azulado.
Vientre y subcaudales verde-azulado.
Cola larga verde con reflejos azulados.
Pico negro, aserrado; patas y pies gris oscuro/marrones-grisáceas claras.
Iris marrón-rojizo oscuro.
Juvenil más apagado que el adulto.
Se diferencia del B. martii totalmente alopátrico por un plumaje generalmente más verde y apagado, con menos rufo, cola más corta y marcadamente menos graduada, timoneras centrales que se estrechan hasta un extremo romo sin raquetas.
Monotípico.
Estrechamente relacionado con B.Martii, y a menudo tratado como con-espespecífico. En estudios recientes se ha demostrado que las razas propuestas: aeruginosos (Mina Gerais, Rio de Janeiro), septentrionalis (Sao Pablo) y abreui (Paraná) solo se basan en variaciones individuales, sin significación geográfica.
Se considera el nombre berlai (Goiás) como inválido (Snow & Kirwan, 2020).
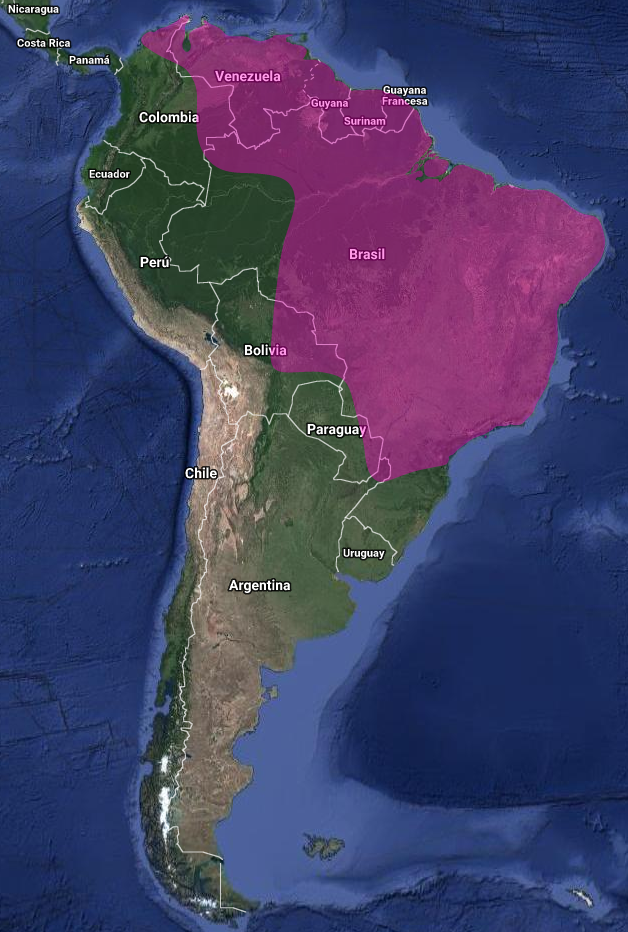
El Yeruvá (Baryphthengus ruficapillus), es una especie endémica de la Mata Atlántica, y anteriormente considerado congénere del Momoto Yerubá occidental (Baryphthengus martii) de la Amazonía occidental y Centroamérica. No obstante, se diferencia claramente por sus partes inferiores mayoritariamente verdosas, interrumpidas sólo por una banda rufa en la parte inferior del pecho; por la corona rufa bien definida, que no se extiende hasta la nuca; y por la ausencia de raquetas evidentes en la cola. La especie se distribuye desde el este de Brasil hasta el sur y este de Paraguay y el extremo noreste de Argentina, habitando bosques de tierras bajas y bosques secundarios de altura, al menos hasta los 1200 msnm (Snow & Kirwan, 2020).
Nombres comunes: Yeruvá (Argentina, Bolivia y Paraguay); Taragón rojizo (Honduras); Burgo rojizo, Momoto yeruvá oriental; Jeruva, Juruva, Juruva-verde; (Brasil) Bururuk, (Botocudo), Jeruva, Pururu, Formigão (Bahía), Juruva-verde? En Goiás es simpátrico con Momotus momota (Burgo) (Sick, 1997:477); En guaraní: «Jiru, Marakana yvyguy», Yirú, marakaná-ihvihgwíh, guirá-hú-tacuara, güirá-payé, güirá-ivigüí, güirá-hú-ivigüí, yakú-tacuara o loro-ybuibui, yeruvá o lorito güí-güí”. Otros: Burgo cola recta, maracaná del suelo, papagayo de la barranca, tutú (de la Peña, 2020:24; Chébez, 1996:137).
En otros idiomas
Breves apuntes históricos:
Apuntamientos para la Historia Natural de las Paxaros del Paraguay y Rio de la Plata. Azara, Félix de. Madrid, 1802-1805. Vol I: 243. (Tutu)
Orden Coraciiformes: 6 familias: En Misiones 2 familias (Alcediniidae y Momotidae). Desde un punto de vista taxonómico, la familia Momotidae comprende seis géneros y catorce especies (del Hoyo, 2020:302); En Argentina se registran dos especies de Momotidae, una de ellas presente en la provincia de Misiones:
Los momótidos (Momotidae), conocidos comúnmente como momotos, barranqueros o guardabarrancos, constituyen una familia de aves tropicales perteneciente al orden Coraciiformes, el cual incluye también a los martines pescadores, los abejarucos y las carracas. Se trata de aves de tamaño mediano que habitan principalmente en selvas densas y cuya distribución se restringe a la región tropical del continente americano. Presentan un plumaje de textura blanda y una cola larga, característica que suelen mover lateralmente de manera frecuente (Sick, 1997:476).
Desde una perspectiva histórica y paleontológica, los momótidos se consideran aves forestales de la región neotropical que se habrían extendido desde Centroamérica. La evidencia fósil incluye un ejemplar del Mioceno (Terciario Superior) hallado en Florida (Becker, 1986), así como restos del Pleistoceno procedentes de las lapas de Escravana, Marinho, Sumidouro y Capão Seco, en el estado de Minas Gerais, datados en aproximadamente 20.000 años antes del presente; dichos registros indican además que la familia estuvo presente en Europa. Filogenéticamente, los Momotidae se encuentran emparentados con los alcedínidos y los trogónidos, relación sustentada en la peculiar morfología del oído interno.
En términos ecológicos y etológicos, los Momotidae son depredadores metódicos de insectos de gran tamaño y pequeños vertebrados en los bosques neotropicales (Winkler et al., 2020). Aunque suelen presentar una coloración llamativa y vocalizaciones frecuentes, pueden resultar sorprendentemente difíciles de localizar, ya que adoptan una estrategia de caza basada en la espera inmóvil de las presas en tránsito. La mayoría de las especies exhiben dos plumas caudales con una forma distintiva de raqueta, la cual se origina cuando las barbas más débiles de los ejes de las plumas se desprenden durante el acicalamiento o como resultado del desgaste normal. Es posible que, como mecanismo de comunicación ante la detección de un depredador, estas aves muevan sus colas características de un lado a otro en ritmos irregulares, deteniendo abruptamente el movimiento en puntos impredecibles del arco de rotación. Asimismo, los Momotidae se encuentran entre los excavadores de nidos y cuidadores más diligentes dentro de los coraciiformes que nidifican en cavidades; sin embargo, pese a esta notable conducta, sus hábitos de nidificación permanecen relativamente poco estudiados (Winkler et al., 2020).
TAXONOMÍA
Los primeros estudios de carácter morfológico y molecular propusieron que la familia Momotidae constituye el grupo hermano de Todidae, y que ambas familias, consideradas conjuntamente, son a su vez hermanas de Alcedinidae (Cracraft, 1981:702/708; Johansson et al., 2001:49). No obstante, análisis morfológicos posteriores situaron a los Momotidae fuera del denominado grupo de los martines pescadores (Livezey & Zusi, 2007). En contraste, investigaciones moleculares más recientes ubican de manera consistente a Momotidae como grupo hermano de Alcedinidae, y a estas dos familias, en conjunto, como hermanas de Todidae. Asimismo, se considera más probable que este clado integrado por tres familias sea hermano del clado conformado por Coraciidae y Brachypteraciidae, y que las cinco familias reunidas mantengan una relación de grupo hermano con Meropidae (Ericson et al., 2006:544; Hackett et al., 2008:1765).
Los géneros y especies reconocidos son los siguientes:
Hylomanes
Hylomanes momotula
Aspatha
Aspatha gularis
Momotus
Momotus mexicanus
Momotus coeruliceps
Momotus lessonii
Momotus subrufescens
Momotus bahamensis
Momotus momota
Momotus aequatorialis
Baryphthengus
Baryphthengus martii
Baryphthengus ruficapillus
Electron
Electron carinatum
Electron platyrhynchum
Eumomota
Eumomota superciliosa
DESCRIPCIÓN
No presentan diferencias externas apreciables entre los sexos. Se caracterizan por un plumaje vistoso y profusamente colorido, así como por un pico fuerte, curvo, relativamente largo, fuerte, algo elevado y ancho en la base, con los lados comprimidos y los márgenes laterales aserrados; estas características varían entre las distintas especies. La lengua es relativamente larga. Las narinas son basales y laterales, con una abertura pequeña y de contorno redondeado. Las alas son relativamente cortas y de forma redondeada, mientras que la cola es larga y graduada, con las dos rectrices centrales generalmente más prolongadas que las restantes. Las patas son cortas, con pies sindáctilos poco especializados. Los dedos son desiguales: el externo es casi tan largo como el medio y se encuentra unido a la base a cierta distancia del segundo; el dedo interno es corto y el posterior, asimismo, corto y débil (Sick, 1997:476).
Una de las características más conspicuas de varias especies es la presencia de “espátulas” en los extremos de las timoneras centrales alargadas, rasgo que contribuye de manera significativa a su atractivo estético. No obstante, la relevancia principal de estas estructuras se vincula con el comportamiento, ya que cumplen una función señalizadora. La génesis de dichas “espátulas” ha sido objeto de debate, y se han propuesto diversas hipótesis al respecto. La mayoría de los autores ha sostenido que las aves arrancan activamente los segmentos correspondientes de las plumas. En este sentido, Ihering (1967) afirmó: “La cola tiene una pequeña sección subapical desprovista de púas, que, dicho sea de paso, son cortadas por la propia ave, que así se adorna. Este hecho constituye un ejemplo quizás único entre las aves”. Sin embargo, esta interpretación no resulta aceptable.
Las plumas implicadas se desarrollan normalmente hasta alcanzar, o casi alcanzar, su longitud definitiva; hasta ese momento no presentan defectos cercanos al ápice, sino únicamente una leve “estrangulación” en la región distal del vexilo, en el punto donde posteriormente se abrirá el espacio que dará lugar a la “espátula”. Este fenómeno ocurre cuando la pluma alcanza cierta edad. Las púas (ramas) localizadas en la zona destinada a desprenderse se tornan quebradizas en la base, adquiriendo un aspecto dentado, y en poco tiempo se desprenden casi espontáneamente o como resultado de una fricción mínima con la vegetación o con las paredes del nido. La extensión del espacio resultante es constante y se encuentra preformada en la estructura de la pluma, tal como se verificó en el material depositado en el Museo de Berlín en 1933. Esta preformación garantiza la integridad de la espátula, que conserva siempre el mismo tamaño; en consecuencia, no son válidas las hipótesis que atribuyen su formación a un trabajo activo del ave o a la influencia de las dimensiones del nido. Además, algunos representantes, como Baryphthengus ruficapillus del sureste de Brasil, que comparte hábitos similares, no desarrollan espátulas y mantienen las rectrices centrales intactas, al igual que el resto de la cola, lo que sugiere la conservación de una etapa evolutiva menos avanzada (Sick, 1997:476).
ALIMENTACIÓN
Presentan una dieta oportunista y variada, basada principalmente en la captura de presas pequeñas. La mayoría de los Momotidae se alimenta fundamentalmente de grandes artrópodos, que suelen capturar mediante salidas cortas desde una percha estacionaria; de manera ocasional, incorporan a su dieta pequeños vertebrados, cangrejos, peces y frutos (Winkler et al., 2020).
Asimismo, estos pájaros recolectan artrópodos posados en hojas y ramas durante el vuelo, de forma similar a los Bucconidae y Galbulidae, y capturan mariposas. Con frecuencia acompañan a las hormigas guerreras, aprovechando los animales que estas ahuyentan, como escarabajos, orugas y ciempiés, e incluso, en ocasiones, pequeños mamíferos, polluelos de aves o lagartijas. También descienden al suelo, donde remueven ramas en descomposición y hojarasca, conducta que explica su denominación de “gallo del bosque”. Para someter a sus presas, las golpean contra el sustrato antes de ingerirlas (Sick, 1997:476).
COMPORTAMIENTO
Los momotos son aves de hábitos arborícolas que se presentan generalmente en forma solitaria o en parejas y mantienen una estrecha asociación con la vegetación alta, tanto arborescente como arbustiva, de la cual rara vez se alejan. Su comportamiento es marcadamente pasivo: permanecen posados en posición vertical, inmóviles y silenciosos durante largos períodos, lo que dificulta su detección cuando no realizan movimientos caudales. Durante estos intervalos, efectúan movimientos pendulares de la cola, tras los cuales se lanzan para capturar presas tales como escarabajos y otros insectos, arañas, lagartijas o pequeñas serpientes en el suelo, así como mariposas, abejas o libélulas en vuelo (Sick, 1997:476).
El vuelo es lento y ondulado, y se ha señalado la existencia de una ligera diferencia en la longitud de la cola entre machos y hembras, siendo esta algo mayor en los primeros, lo que podría cumplir una función de diferenciación sexual. Estas aves suelen posarse a baja altura y, al igual que los Galbulidae, pueden pasar desapercibidas durante períodos prolongados si no mueven la cola. La cola constituye un indicador relevante del estado emocional, con movimientos laterales y verticales abruptos. En el género Baryphthengus, la cola puede elevarse hasta la altura de la coronilla, destacándose las oscilaciones pendulares que les han valido el apelativo de “ave de péndulo”. Resulta particularmente notable la capacidad de detener la cola en cualquier posición, efecto visual que se ve acentuado por la presencia de espátulas. Se han registrado baños de polvo en el Baryphthengus (Sick, 1997:476).
VOCALIZACIONES
El canto de los momotos consiste en un grito bajo y melodioso, emitido principalmente durante la madrugada (suelen iniciarse aún en la oscuridad y cesar antes del alba) y el crepúsculo, que con frecuencia constituye la única evidencia de su presencia. El timbre recuerda al de una paloma o una rana, y es habitual que los miembros de una pareja mantengan una “conversación” mediante llamadas similares. En numerosas ocasiones, la voz representa el único indicio de la presencia de especies como Baryphthengus ruficapillus (Sick, 1997:476).
HÁBITAT
Al igual que la mayoría de los Coraciiformes, los momotos habitan preferentemente los bordes de bosques húmedos y secos, así como áreas semiabiertas e incluso ambientes antropizados, como jardines. En términos generales, los Momotidae se distribuyen en la mayor parte de los hábitats boscosos y arbolados dentro de su área de distribución, y requieren la presencia de taludes o bancos de tierra para la excavación de las cavidades destinadas a la nidificación (Winkler et al., 2020).
REPRODUCCIÓN
Los momótidos nidifican en cavidades excavadas en taludes de tierra o aprovechan madrigueras abandonadas por otros animales, las cuales suelen profundizar. La excavación del nido es realizada por ambos progenitores, quienes trabajan de manera conjunta removiendo el sustrato con las patas y, en ocasiones, con el pico. Como ocurre en la mayoría de los Coraciiformes, la nidificación se lleva a cabo en túneles abiertos en lomas o barrancos, con galerías que pueden variar aproximadamente entre 60 cm y 2 m de longitud, e incluso alcanzar mayor profundidad. La entrada de la madriguera suele ser más ancha que alta y la cámara de incubación se sitúa al final del túnel. Esta estrategia dificulta el acceso al nido por parte de intrusos y depredadores (Sick, 1997:476; Winkler et al., 2020).
Desde el punto de vista reproductivo, las Momotidae son monógamas y, al menos en algunas especies, forman vínculos de pareja duraderos. El tamaño de la puesta es generalmente de cuatro huevos, redondeados, de color blanco brillante, depositados directamente sobre el sustrato del fondo de la cámara de incubación. Ambos padres participan en la incubación y en la alimentación de las crías. El período de incubación se extiende, según las especies y las regiones, entre 17 y 22 días, durante los cuales los progenitores se alternan con escasos relevos diarios (Sick, 1997:476; Winkler et al., 2020).
Crías altriciales y, salvo en el género Aspatha, no desarrollan plumón, sino que les crecen directamente plumas similares a las de los adultos. Los ojos se abren aproximadamente a las dos semanas de edad; no obstante, el crecimiento es rápido y los juveniles abandonan el nido en poco más de tres semanas, o alrededor de un mes tras la eclosión. A pesar de las condiciones insalubres del nido, que suele acumular restos orgánicos y larvas de insectos, los pichones emergen con un plumaje bien desarrollado. En general, las especies parecen reproducirse una sola vez al año y utilizan las madrigueras como refugio únicamente durante la temporada reproductiva. Debido a la profundidad de los nidos y a la escasa frecuencia de cambios en su estructura, muchas especies de Momotidae continúan siendo poco conocidas desde el punto de vista de su biología reproductiva (Sick, 1997:476; Winkler et al., 2020).
ESTADO DE CONSERVACIÓN
La pérdida de hábitat constituye la principal causa por la cual una especie de Momotidae (7 %) se encuentra en categoría de amenaza. En particular, el momoto pico de quilla (Electron carinatum) presenta una distribución restringida en Centroamérica, con baja densidad poblacional y una marcada dependencia del bosque primario, un ambiente que se encuentra en rápido proceso de desaparición; por ello, la especie está clasificada como Vulnerable (Winkler et al., 2020).
Administración de Parques Nacionales (SIB). (2025). Ficha: Coccyzus euleri; recuperado en diciembre de 2025 en https://sib.gob.ar/especies/coccyzus-euleri
Allen, J. A. (1876). List of birds collected by Mr. Charles Linden, near Santarem, Brazil. Bulletin of the Essex Institute, 8: 78–83. (enlace)
Allen, J. A. (1893). On a Collection of Birds from Chapada, Mato Grosso, Brazil by Herbert H. Smith. Part III. Pipridae to Rheidae. Bull. Amer. Mus. Nat. Hist., 5 (10): 107-158. (enlace)
Avibase (s.f.). Ficha: Cuclillo ventriblanco (Coccyzus euleri, Cabanis 1873); Recuperado en diciembre 2025 de https://avibase.bsc-eoc.org/species.jsp?avibaseid=BBA3CB9AC0CFD95D
Banks, R. C. (1988). An old record of the Pearly-breasted Cuckoo in North America and a nomenclatural critique. Bulletin of the British Ornithologists’ Club, 108 (2): 87–91.
Barnett, J. M. & M. Pearman (2001:45). Lista comentada de las aves argentinas. Lynx Edicions, Barcelona. 164 pp. ISBN 84-87334-32-6
Bastos, D. F., R. A. T. Souza, J. Zina & C. A. da Rosa (2019). Seasonal and spatial variation of road-killed vertebrates on BR-330, southwest Bahia, Brazil. Oecologia Australis, 23: 388–402. (enlace)
Bencke, G. A. (2010). New and significant bird records from Rio Grande do Sul, with comments on biogeography and conservation of the southern Brazilian avifauna. Iheringia, Serie Zoologia 100 (4): 391–402. (enlace)
BirdLife International (2024). Species factsheet: Pearly-breasted Cuckoo Coccyzus euleri. Downloaded from https://datazone.birdlife.org/species/factsheet/pearly-breasted-cuckoo-coccyzus-euleri on 03/12/2025
Bodrati, A. (2005). Nuevos aportes a la distribución de algunas especies de aves argentinas. Nuestras Aves, 50: 30-33. (enlace)
Bodrati, A., Cockle, K., Matuchaka, V. & C. Maders (2007). Reserva de la Biósfera Yabotí. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 300-302. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires. (enlace)
Bodrati, A., K. Cockle, J. M. Segovia, I. Roesler, J. I. Areta & E. Jordan (2010). La avifauna del Parque Provincial Cruce Caballero, Provincia de Misiones, Argentina. Cotinga, 32: 41-64. (enlace)
Bodrati, A., J. I. Areta & E. White (2012). La avifauna de la Posada y Reserva Puerto Bemberg, Misiones, Argentina. Nuestras Aves, 57: 63-80 (enlace)
Braun, M. J., D. W. Finch, M. B. Robbins & B. K. Schmidt (2007). A Field Checklist of the Birds of Guyana. 2nd Edition. Publication 121. Biological Diversity of the Guianas Program, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA. (enlace)
Brito, G. R. R., G. M. Kirwan, C. P. Assis, D. H. Firme, D. M. Figueira, N. Buainain & M. A. Raposo (2016). A collection of birds from Presidente Kennedy and adjacent areas, Tocantins: a further contribution to knowledge of Amazonian avifauna between the Araguaia and Tocantins Rivers. Revista Brasileira de Ornitologia, 24 (2):168–184. (enlace)
Cabanis, J. (1873). Protokoll der XLVII Monats-Sitzung. Journal für Ornithologie 21:70–73. (enlace)
Campos Cedeño, E. & A. C. Vallely (2015). First modern North American record and first indication of breeding by Pearly-breasted Cuckoo (Coccyzus euleri) in Panama. North American Birds, 68 (3):302–304.
Capper, D. R., R. P. Clay, N. A. Madroño, B. J. Mazar, I. J. Burfield, E. Z. Esquivel, C. P. Kennedy, M. Perrens & R. G. Pople (2001). First records, noteworthy observations and new distributional data for birds in Paraguay. Bulletin of the British Ornithologists’ Club, 121: 23–37. (enlace)
Carriker, M. A. (1955). Notes on the occurrence and distribution of certain species of Colombian birds. Novedadas Colombianas, 2: 48-64. (enlace)
Cavarzere, V., L. F. Silveira, V. R. Tonetti, P. Develey, F. K. Ubaid, L. B. Regalado & L. F. A. Figueiredo (2017). Museum collections indicate bird defaunation in a biodiversity hotspot. Biota Neotropica, 17 (4): e20170404. (enlace)
Cherrie, G. K. (1916). A contribution to the ornithology of the Orinoco region. Museum of the Brooklyn Institute of Arts and Sciences Bulletin. 2: 133–374. (enlace)
Chébez, J. C. (1994). Los que Se Van. Especies Argentinas en Peligro. Editorial Albatros, Buenos Aires, 604 pp.
Chébez, J. C. (1996). Fauna misionera, catálogo sistemático y zoogeográfico de los vertebrados de la provincia de Misiones (Argentina). Monografía No 5. Págs. 320. L. O. L. A. Bs.Aires. 320 pp. ISBN 950-9725-20-X
Claessens, O., F. P. Brammer, T. Deville A. Renaudier (2011). First documented records of Pearly-breasted Cuckoo Coccyzus euleri for French Guiana, and an overlooked specimen from Ecuador. Bulletin of the British Ornithologists’ Club, 131 (2): 128–133. (enlace)
Claessens, O., M. Giraud-Audine, F. Royer & L. Sénécaux (2017). What is the breeding range and breeding season of Pearly-breasted Cuckoo Coccyzus euleri? New records and breeding in French Guiana. Bulletin of the British Ornithologists’ Club, 137 (1): 3–11. (enlace)
Cockle, K. L., A. Bodrati, M. Lammertink, B. B. Bonaparte, C. Ferreira & F. Di Sallo (2016). Predators of bird nest in the Atlantic Forest of Argentina y Paraguay. Wilson Jour. Ornith., 128: 120-131. (enlace)
Cohn-Haft, M., A. Whittaker & P. C. Stouffer (1997). A new look at the “species-poor” central Amazon: The avifauna north of Manaus, Brazil. Ornithological Monographs, 48: 205–235. (enlace)
Contreras, J. R., L. M. Berry, A. O. Contreras, C. C. Bertonatti & E. E. Utges (1990). Atlas Ornitogeográfico de la Provincia del Chaco – República Argentina. Volume 1. No Passeriformes. Cuadernos Técnicos “Félix de Azara” 1.
Contreras, J. R., F. Agnolin, Y. E. Davies, I. Godoy, A. Giacchino & E. Ríos (2014). Atlas ornito-geográfico de la provincia de Formosa, República Argentina. 1º Ed. Ciudad Autónoma de Buenos Aires: Fundación de Historia Natural Félix de Azara. 450 p. (enlace)
Contreras, J. R., F. L. Agnolin, J. C. Chébez, Y. E. Davies, I. N. Godoy & E. Ríos (MS). Atlas Ornitogeográfico de la Provincia de Misiones, República Argentina. (Inédito).
de la Peña M. R. (1999). Aves Argentinas. Lista y Distribución. Ed. L.O.L.A. (Literature of Latin America). Monografía N.º 18; 244 pp. Buenos Aires. ISSN 0328-1620
de la Peña, M. R. (2015). Aves Argentinas. Incluye nidos y huevos. 1º Ed. Santa Fe: Ediciones UNL; CABA: EUDEBA; Vol 1; 496 pp. ISBN 978-987-657-986-5
de la Peña, M. R. & S. Salvador (2016) Aves argentinas: descripción, comportamiento, reproducción y distribución. Ciconiidae a Heliornithidae. Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie) Año 2016 | Vol. 20 | N° 2 | Pág. 1-627 | ISSN 0325-3856
de la Peña, M. R. (2020). Aves Argentinas. Descripción, Comportamiento, Reproducción y Distribución. (Actualización). Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie), Vol 6:1-348. ISSN 0325-3856
De Oliveria, N. L. da S. & F. R. Dario (2018). Diversity of birds and their ecological interactions in the Mura Indigenous Territory, Brazilian Amazon Rainforest. World News of Natural Sciences, 20: 85–102. (enlace)
del Hoyo, J. (2020). All the Birds of the World. Lynx Edicions, Barcelona. 967 pp. ISBN 978-84-16728-37-4
Di Giacomo, A. G. (2005). Aves de la Reserva El Bagual. En Historia Natural y Paisaje de la Reserva El Bagual, Provincial de Formosa, Argentina. Inventario de la Vertebrados y de la Flora Vascular de un Area Protegida del Chaco Húmedo. Temas de Naturaleza y Conservación, 4 (A. G. Di Giacomo and S. F. Krapovickas, Editors), Aves Argentina/Asociación Ornithológica del Plata, Buenos Aires. pp. 201–465 (enlace)
Dubs, B. (1992). Birds of Southwestern Brazil. Catalogue and Guide to the Birds of the Pantanal of Mato Grosso and its Border Areas. Bertrona Verlag, Küstnacht, Suiza, pp. 1-164, pls. 1-58. (enlace)
EcoRegistros (2025). Cuclillo Ceniciento (Coccyzus euleri). Accedido de https://www.ecoregistros.org el 04/12/2025.
Erritzøe, J., C. F. Mann, F. P. Brammer & R. A. Fuller (2012). Cuckoos of the World. Christopher Helm, London, UK.
Esquivel, A. & S. J. Peris (2011). Aves de San Rafael. Universidad de Salamanca/Asociación Pro Cordillera San Rafael, Paraguay.
Esquivel, A., R. Zarza, R. Tiffer-Sotomayor, A. Díaz, D. Pérez & M. Velázquez (2019). Conservation status and challenges of the Atlantic Forest birds of Paraguay. Diversity, 11: 247 (enlace)
Fernandes-Ferreira, H., S. V. Mendonça, C. Albano, F. S. Ferreira & R. R. N. Alves (2012). Hunting, use and conservation of birds in Northeast Brazil. Biodiversity and Conservation, 21 (1): 221-244. (enlace)
Freile, J. F., R. Ahlman, D. M. Brinkuizen, P. J. Greenfield, A. Solano-Ugalde, L. Navarrete & R. S. Ridgely (2013). Rare birds in Ecuador: First annual report of the Committee of Ecuadorian Records in Ornithology (CERO). Avances, 5 (2): B24–B41. (enlace)
Greenway, J. C. (1978). Type specimens of birds in the American Museum of Natural History, Part 2. Bulletin of the American Museum of Natural History, 161 (1): 1‒305. (enlace)
Griscom, L. & J. C. Greenway (1941). Birds of Lower Amazonia. Bulletin of the Museum of Comparative Zoology at Harvard College, 88: 81–344. (enlace)
Haverschmidt, F. (1968). Birds of Suriname. Oliver & Boyd. Edinburgh and London. Tweedddale Court. Edinburgh. (enlace)
Hellmayr, C. E. (1913). Critical notes on the types of little-known species of neotropical birds, 2. Novitates Zoologicae, 20: 227-256. (enlace)
Hellmayr, C. E. (1929). A contribution to the ornithology of northeastern Brazil. Publications 255 – Field Museum of Natural History, Zoological Series, 12 (18): 235–501. (enlace)
Hilty, S. L. (2003). Birds of Venezuela. (2º Ed.). Princeton University Press, Princeton, NJ, USA. ISBN 0-691-09250-8
Hilty, S. L. & W. L. Brown (1986). A Guide to the Birds of Colombia. Princeton University Press, Princeton, NJ, USA.
Hughes, J. M. (2006). Phylogeny of the cuckoo genus Coccyzus (Aves: Cuculidae): A test of monophyly. Systematics and Biodiversity, 4 (4): 483–488. (enlace)
Hughes, J. M. (2022). Pearly-breasted Cuckoo (Coccyzus euleri), version 2.0. In Birds of the World (S. M. Billerman, Editor). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.pebcuc1.02
International Commission of Zoological Nomenclature (1992). Opinion 1688. Coccyzus euleri Cabanis, 1873 (Aves, Cuculiformes): specific name conserved. Bulletin of Zoological Nomenclature, 49: 178–179. (enlace)
Lawrence, G. N. (1864). Catalogue of birds collected at the island of Sombrero, WI, with observations by AA Julien. Annals of the lyceum of Natural History, New York, 8: 92-106. (enlace1) (enlace2)
Madroño A. N., R. P. Clay, M. B. Robbins, N. H. Rice, R. C. Faucett & J. C. Lowen (1997). An avifaunal survey of the vanishing interior Atlantic forest of San Rafael National Park, Departments Itapúa/Caazapá, Paraguay. Cotinga, 7: 45–53.
Magalhães, J. C. R. (1999). As Aves na Fazenda Barreiro Rico. Plêiade, São Paulo. 215 pp.
Mahler, I.K.F., A. E. Glayson & G. N. Mauricio (2005). A avifauna do Parque Estadual do Turvo, Río Grande do Sul. XI Reunión Argentina de Ornitología, pp. 123. Presentación Oral. (página 130 pdf) (enlace)
Mallet-Rodrigues, F., R. Parrini & B. Rennó (2015). Bird species richness and composition along three elevational gradients in southeastern Brazil. Atualidades Ornitológicas, 188: 39–58. (enlace)
Meyer de Schauensee, R. (1970). A Guide to Birds of South America. Academy of Natural Sciences, Philadelphia, Pennsylvania, USA. (enlace)
Morante-Filho, J. C., D. Faria, E. Mariano-Neto & J. Rhodes (2015). Birds in anthropogenic landscapes: The responses of ecological groups to forest loss in the Brazilian Atlantic forest. PLOS ONE 10(6):e0128923 (enlace)
Moschione, F. & L. Bishels (2005). Listado de las aves del parque Provincial Loro hablador, Provincia del Chaco. Informe técnico proyecto Elé/DFS.
Narosky, T. & D. Yzurieta (2010). Birds of Argentina and Uruguay: a field guide. Vázquez Mazzini Editores, Buenos Aires, Argentina. ISBN 950-99063-6-0
Olrog, C. Ch. (1979). Nueva Lista de la Avifauna Argentina. Ópera Lilloana XXVII. Ministerio de Cultura y Educación. Fundación Miguel Lillo. Tucumán, Argentina. 324 pp. ISSN 0078-5245
Partridge, W. H. (1961). Aves de Misiones nuevas para la Argentina. Neotrópica, 7 (22): 25-28.
Payne, R. B. (2005). The Cuckoos. Oxford University Press, Oxford, UK.
Peters, J. L. (1940). Check-list of Birds of the World. Volume 4. Harvard University Press, Cambridge, MA, USA. (enlace)
Pimentel, L.& F. Olmos (2010). The birds of Reserva Ecológica Guapiaçu (REGUA), Rio de Janeiro, Brazil. Cotinga, 33: 6–22.
Pinheiro, R. T., T. Dornas, E. Santos Reis, M. O. Barbosa & D. Rodello (2008). Birds of the urban area of Palmas, TO: composition and conservation. Revista Brasileira de Ornitologia, 16 (4): 339-347. (enlace)
Pinto, O. M. O. (1978). Novo Catálogo das Aves do Brasil. Primeira parte. Aves não Passeriformes e Passeriformes não Oscines, com exclusão da família Tyrannidae. Conselho Nacional de Desenvolvimento Científico e Tecnológico, São Paulo, Brazil. (enlace)
Planqué, R., Vellinga, W. P., Pieterse, S., Jongsma J. & R. de By (Xeno-canto Foundation). Ficha: Cuclillo ventriblanco (Coccyzus euleri, Cabanis 1873); Recuperado en diciembre 2025 de https://xeno-canto.org/species/Coccyzus-euleri
Remsen, J. V. & M. A. Traylor (1989). An Annotated List of the Birds of Bolivia. Buteo Books, Vermillion, SD, USA.
Remsen, J. V., Jr., M. A. Traylor & K. C. Parkes (1986). Range extensions of some Bolivian birds, 2 (Columbidae to Rhinocryptidae). Bulletin of the British Ornithologists’ Club, 106 (1): 22–32. (enlace)
Restall, R., C. Rodner & M. Lentino (2006). Birds of Northern South America: An Identification Guide. Christopher Helm, London, UK.
Ridgely, R. S. & P. J. Greenfield (2001). The Birds of Ecuador. Volumes 1 and 2. Cornell University Press, Ithaca, NY, USA. 1584 pp. ISBN 9780713661613
Ridgley, R. S., D. Agro & L. Joseph (2005). Birds of the Iwokrama Forest. Proceedings of the Academy of Sciences of Philadelphia, 154: 109–121. (enlace)
Ridgway, R. (1916). The Birds of North and Middle America. Part 7. United States National Museum Bulletin 50. (enlace)
Saibene, C. A., Castelino, M. A., Rey, N. R., Herrera, J. & J. Calo (1996). Inventario de las Aves del Parque Nacional “Iguazú”, Misiones, Argentina. Monografía, 9. Ed. L.O.L.A. Literature of Latin America, Buenos Aires. 68 pp. ISSN 0328-1620
Samuels, I, P. Bichier, J. Clark, T. Milleron & B. O’Shea (2014). Noteworthy records and range extensions from the Caura River watershed, Bolívar state, Venezuela. Cotinga, 36: 27–39.
Santos, F. C. V, W. M. S. Souto, A. S. N. Ribeiro, R. F. P. de Lucena & A. Guzzi (2020). Traditional knowledge and perception of birds in the Paranaíba delta environmental protection area, Northeast Brazil. Acta Scientiarum Biological Sciences, 42: e47722. (enlace)
Sclater, P. L. & G. E. Shelley (1891) Catalogue of the birds of the British Museum. Volume 19. British Museum (Natural History), London, United Kingdom. (enlace)
Short, L. L. (1984). Priorities in ornithology: the urgent need for tropical research and researchers. The Auk, 101 (4): 27. (enlace)
Sick, H. (1997). Ornitologia Brasileira. Editora Nova Fronteira, Rio de Janeiro, Brazil. 912 pp. ISBN 85-209-0816-0
Smith, P., & R. P. Clay (2015). New and noteworthy bird records from “Área de Reserva para Parque Nacional” San Rafael. Paraquaria Natural, 3: 22–26. (enlace)
Spaans, A. L., O. H. Ottema & J. J. M. Ribot (2018). Field Guide to the Birds of Suriname. Brill, Leiden, The Netherlands.
Stotz, D. F., J. W. Fitzpatrick, T. A. Parker & D. K. Moskovits (1996). Neotropical Birds: Ecology and Conservation. University of Chicago Press, Chicago, IL, USA.
Strewe, R. & C. Navarro (2004). New and noteworthy records of birds from the Sierra Nevada de Santa Marta region, north-eastern Colombia. Bulletin of the British Ornithologists’ Club, 124 (1): 38–51. (enlace)
Tonetti, V. R., M. A. Rego, A. C. De Luca, P. F. Develey, F. Schunck & L. F. Silveira (2017). Historical knowledge, richness and relative representativeness of the avifauna of the largest native urban rainforest in the world. Zoologia, 34: e13728. (enlace)
Vidoz, J. Q., A. E. Jahn & A. M. Mamani (2010). The avifauna of Estación Biológica Caparú, Bolivia. Cotinga, 32: 51–68. (enlace)
Vitorino, B. D., A. V. B. Frota, S. K. I. Castrillon & J. R. S. Nunes (2018). Birds of Estação Ecológica da Serra das Araras, state of Mato Grosso, Brazil: Additions and review. Check List, 14: 893–922. (enlace)
Willis, E. O. & Y. Oniki (1990:). Coccyzus euleri (Cabanis, 1873 (Aves, Cuculiformes): proposed conservation of the specific name. The Bulletin of Zoological Nomenclature, 47: 195-197. (enlace)
ESTADO DE CONSERVACIÓN SEGÚN BIRDLIFE INTERNACIONAL: PREOCUPACIÓN MENOR
El Cuclillo ceniciento (Coccyzus euleri) es considerado una especie poco común o rara en la mayor parte de su área de distribución. Su abundancia real sigue siendo insuficientemente conocida, debido tanto a su similitud con el migratorio Coccyzus americanus como a su conducta esquiva y la dificultad de su detección. Además, sus poblaciones pueden experimentar fluctuaciones interanuales asociadas, posiblemente, a la disponibilidad estacional de presas. Aunque la especie está catalogada oficialmente como estable, depende de biomas forestales altamente amenazados, por lo que su situación podría ser más vulnerable de lo que actualmente se reconoce. El tamaño global de la población no ha sido cuantificado, pero se describe a esta especie como “rara” (Stotz y col. 1996).
En Argentina: AvA-SADS-NA: No amenazada según AvA SAyDS 2008 (Aves Argentinas y Secretaría de Ambiente y Desarrollo Sustentable). López-Lanús, B., P. Grilli, E. Coconier, A. Di Giacomo y R. Banchs. Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas /AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina 1º ed. AOP: Buenos Aires, Argentina Res 348 SA-NA: No Amenazada según SAyDS Res 348 (Secretaría de Ambiente y Desarrollo Sustentable). Categorización de aves autóctonas de Argentina. Lista de aves no registradas, endémicas y exóticas de Argentina 2010.
InfoNatura: Animals and Ecosystems of Latin America 2007
El Cuclillo ceniciento (Coccyzus euleri) se distribuye principalmente en Sudamérica al este de los Andes; sin embargo, persisten lagunas importantes respecto de su área reproductiva y su distribución estacional. Esta incertidumbre deriva, en parte, de su marcada similitud morfológica con el Coccyzus americanus (Cuclillo de pico amarillo), especie que también se presenta en el norte y centro de Sudamérica durante la migración boreal y el invierno (Hughes, 2020). La confusión histórica se intensifica por el hecho de que el C. euleri fue considerado durante mucho tiempo una subespecie del C. americanus, lo que vuelve poco informativos numerosos registros antiguos (Erritzøe et al., 2012:295; Barnett & Pearman, 2001:45).
La distribución del C. euleri continúa siendo muy deficiente y sujeta a interpretaciones contradictorias. La propuesta clásica de Meyer De Schauensee (1971:112) que incluye Guyana (Braun et al., 2007:14), Surinam, Venezuela (Mérida, norte de Bolívar y Amazonas), el este de Colombia hasta Cartagena, y el este de Brasil desde Pará hasta Río de Janeiro y Mato Grosso, así como la provincia argentina de Misiones, resulta de extrapolaciones basadas en apariciones esporádicas, sin considerar que la especie alcanza algunas de estas regiones sólo de manera ocasional. Así, en Colombia ha sido sugerida como un probable migrante austral (Hilty & Brown, 1986:217); en Surinam es un migrante poco común entre julio y septiembre, conocido sólo por tres ejemplares históricos (Spaans et al., 2018, citado en Hughes, 2022). Se la describe como “…a rare bird, perhaps only a migrant…” (Haverschmidt, 1968:150); y en Venezuela su presencia es muy localizada, restringida a pocas áreas geográficas y sin claridad sobre su residencia (Meyer De Schauensee & Phelps, 1978:111; Hilty, 2003:350; Payne, 2005:346). En este país se la considera errática y local, con fluctuaciones marcadas en la abundancia interanual, posiblemente relacionadas con brotes de orugas (P. Schwartz).
En Brasil, Sick (1986:319) proporciona información imprecisa “…Rio de Janeiro, Paraná, Mato Grosso, etc…”; en realidad, para Mato Grosso el único antecedente es el registro histórico de Allen (1893), no confirmado posteriormente (Dubs, 1992:47). Pinto (1978:155) ofrece datos puntuales para el Brasil central (Mato Grosso) y oriental (Pará a Paraná), sugiriendo cierta continuidad poblacional en el centro-este del país. Es probable que algunas de las poblaciones dispersas de Venezuela y del centro-este de Brasil sean reproductivas, y que la especie exhiba un grado elevado de trashumancia. Este patrón, sumado a la escasez de observaciones y colecciones en un subcontinente con baja densidad de ornitólogos (Short, 1984:892), dificulta la construcción de un marco biogeográfico coherente.
Los registros específicos disponibles refuerzan este carácter fragmentario. En Colombia se la considera rara en el norte y oriente hasta los 400 m (enero), con observaciones en la costa caribeña cerca de Cartagena, en el sureste de Guainía a lo largo del Río Negro (Macacuní; Hilty & Brown, 1986:217), y en mayo en el Parque Nacional Tayrona, Neguanje y la Reserva Natural Los Besotes (Strewe & Navarro, 2004:41). En Venezuela se documenta de manera errática en Mérida, Distrito Federal (San José de Los Caracas), sureste de Apure (Puerto Páez), norte de Bolívar (Caicara, alto Río Caura, Ciudad Bolívar, Upata, Guasipati) y oeste de Amazonas (Caño Cuao, El Carmen), además de Isla Margarita (Hilty, 2003:350), con registros adicionales en el río Nichare (abril) y el bajo Caroní cerca de Santa Elena (Samuels et al., 2014, citado en Hughes, 2022).
En la Guayana Francesa la especie es rara, con ocho registros que totalizan trece individuos entre 2009 y 2015, principalmente en agosto y septiembre (Claessens et al., 2011:129; Claessens et al., 2017:4). En Ecuador se ha consignado como migrante poco común (marzo; Payne, 2005:346; Freile et al., 2013:B34; Ridgely & Greenfield, 2001).
En Brasil, aunque generalmente rara, podría ser poco común en regiones meridionales (Claessens et al., 2017:7; Payne, 2005:346; De Oliveira et al., 2018:99; Brito et al., 2016:179; Pinheiro et al., 2008:343; Morante-Filho et al., 2015:7; Vitorino et al., 2018:903; Cavarzere et al., 2017:8; Tonetti et al., 2017:12; Pimentel & Olmos, 2010:6; Bencke, 2010; Erritzøe et al., 2012:295).
En Bolivia aparece como migrante poco común en abril, mayo y octubre (Payne, 2005:346; Vidoz & Mamani, 2010; Remsen et al., 1986:25; Remsen & Traylor, 1989). Finalmente, en Paraguay constituye un visitante estival raro entre octubre y noviembre (Esquível & Peris, 2011; Esquível et al., 2019:15; Smith & Clay, 2015:24; Capper et al., 2001:28; Payne, 2005:346).
Se conocen dos registros de C. euleri fuera de su distribución típica en Sudamérica. El primero corresponde a un espécimen inmaduro recolectado en octubre de 1863 en la isla Sombrero, ubicada al este de las Islas Vírgenes y al noroeste de Anguila, en el punto más septentrional de las Antillas Menores (Banks, 1988:90). Este individuo, curiosamente, constituye el espécimen tipo (descrito inicialmente como Coccyzus julieni y posteriormente asignado a Coccyzus euleri, a pesar de proceder de una localidad claramente extralimital. Su hallazgo dio origen a una prolongada controversia sobre la nomenclatura y la distribución geográfica de la especie. El segundo registro corresponde a la observación de una pareja en reproducción en julio en el Lago Bayano, Panamá (Campos-Cedeño & Vallely, 2015, en Hughes, 2022). No obstante, dos supuestos avistamientos adicionales en el centro de Panamá, en junio de 2007 y junio de 2009, fueron rechazados por el Comité de Registros de Panamá (D. Montañez y G. Berguido in litt., en Claessens et al., 2011).
Distribución geográfica en ARGENTINA (de la Peña, 2020:99; de la Peña & Salvador, 2016:338; de la Peña, 2015:303) (de la Peña, 1999:84; EcoRegistros 20 registros en en https://www.ecoregistros.org/ficha/Coccyzus-euleri; eBird 2025, en https://ebird.org/species/pebcuc1/AR-N)
En Argentina, el estatus local de conservación del Coccyzus euleri permanece indefinido (Contreras et al., 2014:281). Es posible que la intensificación reciente de los esfuerzos de observación esté revelando una presencia más frecuente de la especie. No obstante, su situación general podría evidenciar una disminución numérica y cierta retracción areal en sus movimientos migratorios, como sugiere la casi desaparición de la población migrante en Misiones (Contreras et al., Msc), aunque ejemplares han vuelto a registrarse en el Parque Nacional Iguazú (Chébez, 1994:426). Asimismo, la especie ha sido documentada en el Chaco argentino (Bodrati, 2005:31).
Contreras (1990:119) destaca que “…su presencia en el norte argentino, el Paraguay y Bolivia es más probable que lo que los presentes registros hacen suponer. La falta de prospección sistemática y de larga duración y la escasez de estudios regionales no permite, en la región de probable aparición de la especie, especulaciones acerca de su posible presencia.”. En este sentido, la objeción planteada por Chébez (1994:426) respecto de la observación de Canzioni (en Contreras, 1990:119) resulta poco sostenible, ya que presupone una fijación estricta al hábitat que no se ajusta al comportamiento real de la especie. El registro boliviano (Remsen, 1986:25), por ejemplo, procede de un ambiente muy diferente al de Misiones.
Más recientemente, su presencia ha sido confirmada en el Parque Estadual do Turvo (Mahler et al., 2005, en Hughes, 2022), mientras que los registros del estado de Paraná son los únicos claramente afines a los de Misiones.
El Coccyzus euleri se reproduce principalmente en bosques húmedos siempreverdes, bosques de galería y bosques secundarios. Durante la invernada en la Amazonia, utiliza bosques primarios de tierra firme y bosques ribereños dominados por el Ambay (Cecropia). Ocurre típicamente desde el nivel del mar hasta los 900 msnm de altitud (Erritzøe et al., 2012:296).
En Colombia y Venezuela se registra en bosques arenosos, matorrales y bosques de galería (Hilty & Brown, 1986:217); adicionalmente, en Colombia ocupa bosques secos tropicales (Strewe & Navarro, 2004:41) y en Venezuela bosques secundarios (Samuels et al., 2014). En esta última región se encuentra desde los 700 msnm al norte del Orinoco hasta los 350 msnm al sur del río (Hilty, 2003:350).
En Guyana utiliza bosques de tierras bajas (Braun et al., 2007:14), mientras que en Surinam emplea bordes costeros de manglar y bosques de sabana (Spaans et al., 2018, en Hughes, 2022). En la Guayana Francesa ocupa bosques primarios de dosel alto y bordes de bosque, así como bosques secundarios densos de dosel bajo y arboledas de tierras bajas (Claessens et al., 2017:5). En Bolivia ha sido registrada en hábitats de sabana (Vidoz et al., 2010) y en Ecuador en formaciones secundarias de borde (Freile et al., 2013:B24).
En Brasil utiliza el bosque primario de «terra firme» (Cohn-Haft et al., 1997) y el bosque estacional semideciduo (Vitorino et al., 2018:903). Generalmente se distribuye en altitudes de hasta 900 msnm, pero existe un registro en Itatiaia a 1.200 msnm (Mallet-Rodrigues et al., 2015:48). También emplea el bosque atlántico costero del sureste de Brasil, el este de Paraguay y el noreste de Argentina (Madroño et al., 1997). En Panamá se ha observado en bosques latifoliados secundarios (Campos-Cedeño & Vallely, 2015, en Hughes, 2022).
Información adicional señala su presencia en “…capoeira…” (Sick, 1997:387) y en ambientes típicos de la Chapada dos Guimarâes descritos como “…planalto…» de 700 a 800 metros acima do nível do mar, com matas ciliares nos vales e cerrados ou campos com buritis (Mauritia flexousa L.) nos planaltos…” (Willis & Oniki, 1990:21; véase también Allen, 1893 en Contreras, 1990:119). Contreras (1990:119) agrega que en Colombia puede ocupar matorrales y selvas ribereñas.
En Argentina se ha citado en selvas y bosques de transición (de la Peña, 2020:98).
Movimientos y migración
Los movimientos estacionales del Coccyzus euleri son poco conocidos. La especie es, al menos parcialmente, un migrante austral: algunos individuos se reproducen en el sur y el este de Brasil, el norte de Argentina y Paraguay durante el verano austral, y migran luego hacia el norte de Sudamérica en la estación no reproductiva. No obstante, los registros de reproducción al norte del Ecuador y en Panamá, entre enero y agosto, sugieren que ciertas poblaciones septentrionales podrían ser no migratorias (Payne, 2005:346; Claessens et al., 2017:3; Erritzøe et al., 2012:296).
Diversos autores subrayan que es un cuclillo sudamericano poco conocido y aparentemente raro. La posible confusión con el migrante boreal C. americanus, la escasez de registros confiables al norte del Amazonas y el comportamiento esquivo de esta especie forestal dificultan la delimitación precisa de su área de reproducción y su estatus en el norte de Sudamérica. En consecuencia, se la considera preferentemente un migrante austral que se reproduce principalmente en el sur y este de Brasil, el norte de Argentina y Paraguay, y migra posteriormente a la región septentrional del continente (Payne, 2005:346; Claessens et al., 2011:128; Erritzøe et al., 2012:290).
El Cuclillo ceniciento se caracteriza por su comportamiento discreto y esquivo, permaneciendo con frecuencia oculto entre la vegetación (Erritzøe et al., 2012:295), razón por la cual puede pasar fácilmente desapercibido. Un individuo observado forrajeando tranquilamente junto a una pequeña bandada mixta en un bosque seco cercano a Bolívar, Venezuela, permaneció inmóvil entre el follaje durante aproximadamente un minuto antes de emprender un vuelo repentino tras un movimiento ascendente de la cola (Hilty, 2003:350). La especie es típicamente solitaria, aunque ocasionalmente puede registrarse en parejas.
Si bien no suele considerarse una especie particularmente social, ha sido observada integrando bandadas mixtas activas en el dosel en Manaus, Brasil (Cohn-Haft et al., 1997:215), en las proximidades de Bolívar, Venezuela (Hilty, 2003:350), y en Kourou, Guayana Francesa, donde se encontraba asociada con Cotinga cotinga (Cotinga pechimorada), Xipholena punicea (Cotinga púrpura), Polioptila guianensis (Perlita guayanesa) y Cyanicterus cyanicterus (Frutero lamiazul; Claessens et al., 2017:4).
No se ha documentado directamente la depredación de nidos; sin embargo, la pérdida de un nido en la Guayana Francesa podría atribuirse a la acción de un grupo de monos ardilla comunes (Saimiri sciureus), que se encontraban activos en el área en ese momento (Claessens et al., 2017:8).
VOCALIZACIONES
Emite temprano por la mañana durante pocos minutos o en forma muy esporádica hasta las primeras horas de la tarde, siendo esta actividad notable durante pocos días entre los meses de octubre y noviembre
El canto primario del Cuclillo ceniciento, conocido como «Kuoup Call», consiste en una serie de notas fuertes, ásperas y de timbre semejante al de una rana, generalmente entre 5 y 15 emisiones, aunque en ocasiones supera las 20. Estas notas, que suelen producirse a un ritmo aproximado de una por segundo y con desaceleración hacia el final de la secuencia (Hilty, 2003:350), recuerdan a las notas terminales lentas del Cuclillo pico amarillo (Coccyzus americanus); sin embargo, esta última especie es típicamente silenciosa en sus áreas de invernada sudamericanas (Claessens et al., 2017:3). De manera complementaria, la especie emite una llamada de tipo cascabel ascendente, seguida de 4 a 9 notas acentuadas («tuctuctuctuc, tówlp»), similares a las de C. americanus (Hilty & Brown, 1986:217). Aunque el Tingazú (Piaya cayana) también produce un sonido de cascabeleo, carece de las notas terminales acentuadas (Bencke, 2010). El «rattle call» es menos frecuente que el «Kuoup Call» (Hilty & Brown, 1986:217).
Según Claessens et al. (2017:5), antes del inicio de la incubación se registran dos vocalizaciones principales: el canto, emitido por ambos sexos, y la llamada de cascabel, interpretada como señal de contacto o agresión, pues suele generarse en respuesta a la reproducción de vocalizaciones (playback), más que como señal de alarma. La actividad vocal es intensa durante el cortejo y la construcción del nido, pero disminuye marcadamente al comenzar la incubación (Claessens et al., 2017:5). Adicionalmente, responde de manera activa al playback, prolongando su actividad vocal y emitiendo particularmente la llamada de cascabel (Freile et al., 2013:B24; Claessens et al., 2017:4).
Asimismo, se han documentado interacciones vocales entre individuos: en la Guayana Francesa, un miembro de una pareja emitió el «Kuoup Call», mientras el otro respondió con la llamada de cascabel desde una percha ubicada a 5–6 m de altura (Claessens et al., 2017:4). Entre otras descripciones tempranas, Sick (1997:381) caracterizó la voz como ventrílocua (“kjoa”)
ALIMENTACIÓN
La información disponible acerca de la dieta del Cuclillo ceniciento es relativamente limitada. Sin embargo, al igual que muchas especies de la familia Cuculidae, es fundamentalmente insectívoro. Consume diversos invertebrados, con una clara preferencia por las orugas, y busca alimento principalmente en el dosel medio y superior del bosque, así como en zonas de borde forestal (Erritzøe et al., 2012:295). Su comportamiento alimentario es predominantemente solitario, aunque ocasionalmente puede integrarse en bandadas mixtas de especies (Erritzøe et al., 2012:295). En Brasil, se observó un individuo capturando una oruga cuya longitud era aproximadamente 1,5 veces la de su pico (Bencke, 2010).
REPRODUCCIÓN
La distribución reproductiva y los movimientos estacionales del Cuclillo ceniciento permanecen insuficientemente documentados. Tradicionalmente se pensaba que era una especie exclusivamente migratoria, que anidaba en el sur y este de Brasil, el norte de Argentina y Paraguay durante el verano austral, e invernaba al norte del ecuador, desde Colombia hasta la Guayana Francesa. No obstante, registros de reproducción en Colombia (Carriker, 1955:52), Venezuela (Claessens, 2017:7; Cherrie, 1916:311; Erritzøe et al., 2012:296), Guayana Francesa, el norte de Brasil (Claessens, 2017:7; Payne, 2005:346) y Panamá (Campos-Cedeño & Vallely, 2015, citado en Hughes, 2022) entre enero y agosto cuestionan esta interpretación y sugieren que las poblaciones septentrionales podrían ser residentes (Claessens, 2017:10).
El estudio de los registros reproductivos revela dos áreas de reproducción disyuntas: una en el sur de Sudamérica (sur de Brasil, Paraguay -Capper et al., 2001:28- y Argentina) y otra en el norte (este de Panamá, norte de Colombia, Venezuela, extremo norte de Brasil y las Guayanas), con más de la mitad de los registros al norte del ecuador. Resulta notable que la especie haya pasado inadvertida en regiones con comunidades ornitológicas consolidadas, como la Guayana Francesa, donde los primeros registros datan de 2009 (Claessens et al., 2011; Claessens, 2017:7) y Venezuela, donde se considera vagabunda (Hilty, 2003:350). En la Guayana Francesa, los registros confirmados (9 de agosto – 17 de septiembre) coinciden con la condición de migrante austral, lo que respalda la hipótesis de reproducción ocasional fuera del área típica, como sugiere también el registro en Roraima. La abundancia inusualmente alta en 2013 podría indicar una irrupción excepcional que facilitó la reproducción oportunista (Claessens, 2017)
En Argentina, la nidificación ha sido confirmada en Misiones en varias ocasiones. De la Peña (2020:98) menciona nidos localizados en la provincia y señala que la reproducción ocurre en octubre y noviembre. Partridge (1961:25) registró ejemplares con huevos listos para la puesta y testículos activos entre octubre y diciembre en arroyo Urugua-í. Otros nidos fueron documentados en el Parque Nacional Iguazú (Pearman en Bodrati et al., 2012:66), en Posada y Reserva Puerto Bemberg (Bodrati et al., 2012:65) y en el Parque Provincial Cruce Caballero, donde Cockle et al. (2016:122) observaron el 23 de noviembre de 2011 un evento de depredación: un Ramphastos dicolorus espantó a un adulto de C. euleri de un nido en copa situado a ~20 m en un Cedro blanco (Cedrella fissilis) consumiendo al menos un huevo antes de retirarse. El adulto regresó al nido pocos segundos después.
El comportamiento reproductor, incluida la incubación y el cuidado parental, ha sido escasamente observado. La especie es probablemente monógama. Se ha descrito cortejo con alimentación previa a la cópula: el macho ofrece una oruga, la hembra adopta una postura sumisa y posteriormente copulan, aunque no siempre se confirma la aceptación del alimento. La cópula ocurre a veces cerca del nido (≈10 m) y puede ser inducida también por la presentación de material de nidificación (Capper et al., 2001:28; Claessens, 2017:5). Llamativamente, se ha registrado cópula durante la estación no reproductiva tras la reproducción de vocalizaciones (playback), lo que podría indicar una defensa de pareja frente a un rival percibido (R. Czaban in litt. 2014; M. Cohn-Haft in litt. 2014 en Claessens, 2017:9). Este comportamiento sugiere emparejamientos durante la migración o la invernada, fenómeno no descrito previamente en los Cuculidae (Claessens, 2017:9).
Los nidos activos rara vez se detectan debido a su ubicación en vegetación densa (Claessens, 2017:5). Se han registrado entre 3 y 7 m de altura en un Ceibo Erythrina crista-galli en Argentina y en un Palo mulato (Bursera simaruba) en Panamá. En la Guayana Francesa, un nido fue hallado cerca de la punta de una rama horizontal a ~10 m de altura, oculto por lianas y protegido por un sotobosque de palmeras espinosas; el dosel circundante no superaba los 15 m. Un segundo nido, atípico, se ubicó en un bosque bajo (~10 m) sobre una barra arenosa rodeada de manglares, sabanas pantanosas y áreas cultivadas, a menos de 250 m del borde de la sabana (Claessens, 2017:5).
Ambos sexos participan en la construcción del nido. En la Guayana Francesa, el macho recolectó materiales —ramitas, lianas muertas, filamentos similares a hierba, musgo y una hoja seca— y los entregó a la hembra, siguiendo rutas discretas entre el follaje (Claessens, 2017:5).
El nido consiste típicamente en una plataforma suelta de palos delgados, similar al nido del Cuclillo pico amarillo (Campos-Cedeño & Vallely, 2015, citado en Hughes 2022).
Los huevos son elípticos, verde azulado pálido, y la puesta probablemente en número de 2 a 4 (Payne, 2005:347). Se desconoce la duración del periodo de incubación, aunque podría asemejarse a la de otros Coccyzus (9–12 días), con incubación iniciada desde la puesta del primer huevo y eclosión asincrónica. La incubación podría no ser biparental: en un nido de la Guayana Francesa solo se observó a la hembra incubando durante cinco días, adoptando una postura característica con el pecho dentro del nido y la cola elevada; el nido fue abandonado siete días después, posiblemente por depredación (Claessens, 2017:8).
A diferencia de numerosos Cuculidae, el C. euleri no es un parásito de cría obligado, aunque se requieren más estudios para confirmar plenamente este aspecto.
Según los autores (Hughes, 2022; Payne, 2005:345; Erritzøe et al., 2012:295; Hilty, 2003:350; Hilty & Brown, 1986:217; Brito et al., 2016:179)
La especie es sexual y estacionalmente monomórfica.
Adulto: Las partes superiores presentan un tono marrón grisáceo uniforme, con un ligero brillo bronceado. La región facial muestra un gris más oscuro por encima y detrás del ojo, lo que confiere una apariencia semejante a una máscara. El iris varía de marrón a negro. El anillo ocular es gris a negro en la hembra adulta, aunque ocasionalmente puede ser amarillo o rojo.
El pico moderadamente largo y decurvado con la punta ganchuda. El maxilar (mandíbula superior) es predominantemente marrón a negro. La mandíbula inferior y la base del maxilar son amarillas a anaranjadas, con la punta de la mandíbula inferior de color negro. El interior de la cavidad bucal en el adulto es también negro. El iris, el pico y las patas del juvenil son equivalentes a los del adulto.
Las alas son marrón grisáceo, con las secciones internas de las primarias blancas y las cubiertas alares inferiores blanco nacarado.
La cola, larga y graduada, es marrón grisáceo por arriba y negruzca por debajo. Las cubiertas infracaudales presentan un tono gris nacarado claro. La parte inferior de la cola es generalmente negruzca, con grandes manchas blancas y redondeadas en las puntas de las timoneras: T3 (13 mm), T4 (18 mm) y T5 (22 mm). T1 es gris parduzco y carece de punta blanca, mientras que T2 es negruzca con estrechas manchas blancas (3 mm).
Las partes inferiores, desde la garganta hasta el pecho, exhiben un color blanco grisáceo nacarado; el vientre es blanco, mientras que las axilares y las cubiertas infraalares son blanco nacarado.
Los pies zigodáctilos (dos dedos internos dirigidos hacia adelante y dos externos invertidos) presentan coloración gris a gris azulada, con la suela amarillenta y las garras negras.
Juvenil: El juvenil es similar al adulto, aunque presenta bordes finos y pálidos en las alas. Las remeras son más estrechas y puntiagudas, con manchas blanquecinas inferiores menos definidas que gradúan hacia el marrón. La cola carece de manchas en R1; R2 muestra un tenue rastro blanco en la punta; y R3 (8 mm), R4 (15 mm) y R5 (15 mm) presentan manchas blancas. Las cubiertas auriculares no son más oscuras que la corona, por lo que no exhiben la apariencia de máscara facial característica del adulto.
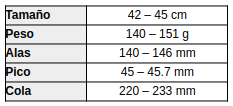
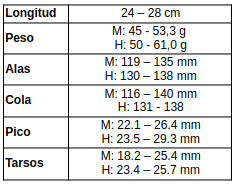
Medidas (Payne, 2005:345; Erritzøe et al., 2012:295; Hilty, 2003:350; Hilty & Brown, 1986:217; Brito et al., 2016:179; de la Peña, 2020:98)
Mudas
Poco conocida. Probablemente muda una vez al año (Payne, 2005:345). Las primarias se mudan siguiendo un patrón de muda irregular, en el que la muda avanza o retrocede saltándose una o más primarias adyacentes (Payne, 2005:345).