- Género Odontophorus
- Odontophorus capueira (Urú)
-
-
Chajá | Bibliografía
- Acevedo, D. & F. Bruno (2007). Lista de Aves del Parque Don Tomás. Santa Rosa, La Pampa. Editorial L. y M. Gral. Acha. La Pampa. (enlace)
- Acevedo, G. S., H. A. Valderrey, M. Cervio, A. Talone, S. Mallet, G. Antenucci & C. J. Rojas (2023). Avifauna de la ciudad de General Pico, La Pampa, Argentina: nuevos registros, distribuciones y evidencias de nidificación para la provincia. Nuestras Aves, 68: 95-117. (enlace)
- Antelo, C. M. & Z. J. B. Fernández (2013). Las aves no passeriformes de Tucumán, Argentina. Miscelánea, 132. Fundación Miguel Lillo, Argentina. (enlace)
- Aravena, R. O. (1928). Notas sobre la alimentación de las aves. Hornero, 4: 153-166 (enlace)
- Avibase (2026). Ficha: Chajá Común (Chauna torquata, Oken, L 1816); recuperado en junio 2026 de https://avibase.bsc-eoc.org/species.jsp?avibaseid=91194BE324AB9F4C
- Azpiroz, A., M. Mattalia, B. López-Lanús & L. L. Sosa (2013). Caracterización ornitológica y productiva del campo “Gral Ávalos” Monte Caseros, Corrientes, informe técnico de Aves Argentinas/AOP: diciembre 2010. En pp. 351-373: G. D. Marino, F. Miñarro, M. E. Zaccagnini y B. López-Lanús (eds.). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. 576 pp. ISBN 978-987-45316-1-2. (enlace)
- Babarskas, M., J. O. Veiga & F. C. Filiberto (1995). Inventario de Aves del Parque Nacional El Rey, Salta, Argentina. Monografía Especial L. O. L. A. Nº 6, 44 Págs. Buenos Aires.
- Babarskas, M., E. Haene & J. Pereira (2003). Aves de la Reserva Natural Otamendi. En Haene, E. y J. Pereira (Eds.). Fauna de Otamendi. Inventario de los animales vertebrados de la Reserva Natural Otamendi, Campana, Buenos Aires, Argentina. Temas de Naturaleza y Conservación. Monografía de aves argentinas 3: 47-113. Aves Argentinas/AOP. Buenos Aires (enlace)
- Barnett, J. M. & M. Pearman (2001). Lista comentada de las aves argentinas. Lynx Edicions, Barcelona. 164 pp. ISBN 84-87334-32-6.
- Barrionuevo, C., D. Ortiz & M. Castillo (2009). Presencia y distribución de algunas aves de ambientes acuáticos de la provincia de Catamarca, Argentina. Xolmis, 4: 27-33.
- Bauni, V., R. E. Carini, S. Bogan, J. M. Meluso, T. N. Abad, M. Homberg & A. Giacchino (2017). Inventario de vertebrados del “Paisaje Protegido Delta Terra”, primera sección de islas del delta de Tigre, provincia de Buenos Aires, Argentina. Nótulas Faunísticas. Segunda Serie, 210: 1-11. (enlace)
- Belton, W. (1984). Birds of Rio Grande do Sul, Brazil. Part 1: Rheidae through Furnariidae. Bulletin of the American Museum of Natural History, 178 (4): 371–631. (enlace)
- Beltzer, A. H. (1981). Lista preliminar de las aves del departamento La Paz (Entre Ríos, Argentina). I. No Passeriformes. Historia Natural, 2: 53-56.
- Beltzer, A. H. (1986). Estudio preliminar de la avifauna de la Laguna del Cristal (Cuenca del río Saladillo, Santa Fe, Argentina). Historia Natural, 8: 65-74.
- Beltzer, A. H., P. A. Collins & M. A. Quiroga (2006). Atlas ornitogeográfico de la provincia de Entre Ríos. Serie Climax No 13. Ceride-Conicet. Santa Fe.
- BirdLife International (2024). Species factsheet: Southern Screamer Chauna torquata. Downloaded from https://datazone.birdlife.org/species/factsheet/southern-screamer-chauna-torquata on 21/06/2026
- Blake, E. R. (1977). Manual of Neotropical Birds. Volume 1: Spheniscidae (Penguins) to Laridae (Gulls and Allies). University of Chicago Press, Chicago, IL, USA. (enlace)
- Bodrati, A., E. Mérida, G. Bodrati & E. Sierra (2006). Avifauna del talar de Vuelta de Obligado y de sus ambientes contigüos. San Pedro, Provincia de Buenos Aires, Argentina. Pp.: 117-124, en: Mérida, E. y J. Athor (Editores). Talares bonaerenses y su conservación. Fundación de Historia Natural “Félix de Azara”. Buenos Aires (enlace)
- Bodrati, A. & A. Pietrek (2000). Relevamiento de los recursos biológicos del Parque Provincial “Pampa del Indio” (Departamento Libertador Gral. San Martín, Chaco). AOP. Inf. Inéd. Buenos Aires.
- Bollero, V. & R. Ramírez (2012). Relevamiento de la avifauna de Laguna El Hinojo, Venado Tuerto, Santa Fe, Argentina. Período 2009/2011. Biológica, 15: 119-126. (enlace)
- Brady, S. (2020). Southern Screamer (Chauna torquata), version 1.0. In Birds of the World (T. S. Schulenberg, Editor). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.souscr1.01
- Brandán, Z. J. & C. I. Navarro (2009). Lista actualizada de las aves de la provincia de Tucumán. Fundación Lillo. Tucumán (enlace)
- Brandolin, P., R. Martori & M. Ávalos (2007). Variaciones temporales de los ensambles de aves de la Reserva Natural de Fauna Laguna La Felipa (Córdoba, Argentina). Hornero, 22 (1): 1-8. (enlace)
- Brumfield, R. T., O. Maillard Z., R. C. Faucett, G. Sánchez, V. G. Rohwer, J. C. Catari, C. G. Schmitt, D. C. Schmitt, R. Strem & A. M. Mamani (2004). Birds of the Laguna Kaucaya area, a semi-humid valley in the Andean foothills of Departamento Santa Cruz, Bolivia. Ornitología Neotropical, 15 (3): 381–398. (enlace)
- Bruno, F., D. Acevedo, J. L. Maceda & M. I. Mero (2007) Reserva Natural Municipal Chadilauquen. Aves. Municipalidad de Embajador Martini. La Pampa (enlace)
- Bruno, G. & E. Vivas (2020). Aves del Mar de Ansenuza: Guía de campo. Ed. Ecoval. Córdoba. 306 pág.
- Bucher, E. H. & G. Herrera (1981). Comunidades de aves acuáticas de la laguna Mar Chiquita (Córdoba, Argentina). Ecosur, 8 (15): 91-120. (enlace)
- Burgos, F. G., J. L. Baldo & F. M. Cornell (2009). Lista de las aves de la Provincia de Jujuy, Argentina. Secretaría de Turismo y Cultura de Jujuy. S. S. de Jujuy. (enlace)
- Camperi, A. R., Darrieu, C. A., & M. Juárez (2008). Avifauna de la provincia de La Rioja (Argentina): lista comentada de especies. Acta zoológica lilloana, 76-97. (enlace)
- Canavelli, S., M. E. Zaccagnini, J. Torresín, N. Calamari , M. de la Paz Duccomun & P. Capllonch (2004). Monitoreo extensivo de aves en el centro-sur de Entre Ríos. Insugeo, Miscelánea, 12: 349-362. (enlace)
- Carboneras, C. (1992). Family Anhimidae (Screamers). Pages 528-535 in J. del Hoyo, A. Elliot, and J. Sargatal (editors). Handbook of the birds of the world. Volume 1. Lynx Edicions, Barcelona, Spain. (enlace)
- Carma, M. I. (2009). Relevamiento de aves de la provincia de Catamarca. Consejo Federal de Inversiones. 1º Ed., Buenos Aires. 500 pp. ISBN 978-987-510-081-7 (enlace)
- Castellanos, A. (1931). Aves del valle de los Reartes (Córdoba). Hornero, 4: 361-391 (enlace)
- Castro, D. del C. & A. C. Cicchino (1992). Algunas especies de Menoponidae (Insecta: Phthiraptera; Amblycera) parásitas de aves en la provincia de Buenos Aires, Argentina. Rev. Asoc. Cienc. Nat. Litoral, 23: 59-69 (enlace)
- Chatellenaz, M. L. (2005). Aves del Valle del Río Paraná en la provincia del Chaco, Argentina: riqueza, historia natural y conservación. Miscelánea, 14: 528. (enlace)
- Chatellenaz, M. L., P. Cano, C. Saibene & H. A. Ball (2010). Inventario de las aves del Parque Nacional Mburucuyá (provincia de Corrientes, Argentina). Acta Zool. Lilloana, 54: 139-160.
- Chébez, J. C. (1996). Fauna misionera, catálogo sistemático y zoogeográfico de los vertebrados de la provincia de Misiones (Argentina). Monografía No 5. Págs. 320. L. O. L. A. Bs.Aires. 320 pp. ISBN 950-9725-20-X
- Chébez, J. C., N. R. Rey, M. Babarskas & A. G. Di Giacomo (1998). Las aves de los Parques Nacionales de la Argentina. Monografía Especial L.O.L.A. No 12, 126 págs. Bs.As.
- Chébez, J. C. (2005). Guía de las Reservas Naturales de la Argentina. (5). Zona Centro. 288 págs. Albatros, Buenos Aires. ISBN-13: 978-950-241060-9
- Coconier, E. G., López Lanús, B., Roesler, I., Moschione, F., Pearman, M., Blendinger, P., Bodrati, A., Monteleone, D., Casañas, H., Pugnali, G. & M. E. Álvarez (2007). Lista comentada de las aves silvestres de la unidad de gestión Acambuco. Pp: 32-103. En Coconier, E. G. (Ed.). Las aves silvestres de Acambuco, Provincia de Salta, Argentina. Temas de Nat. y Cons. No 6. Nuestras Aves A.O.P.
(enlace) - Codesido, M., C. M. González-Fischer & D. N. Bilenca (2013). Landbird assemblages in different agricultural landscapes: a case study in the pampas of central Argentina. Cóndor, 115 (1): 8–16. (enlace)
- Contreras, J. R. (1981). Lista preliminar de la avifauna correntina. I. No Passeriformes. Historia Natural, 2: 21-28.
- Contreras, J. R. (1987). Lista preliminar de la avifauna de la provincia de Formosa, República Argentina. Historia Natural, VII (4): 33-52.
- Contreras, J. R., L. M. Berry, A. O. Contreras, C. C. Bertonatti & E. E. Utges (1990). Atlas ornito-geográfico de la provincia del Chaco. República Argentina. I. No passeriformes. Cuad. Téc. Félix de Azara Nº 1. 164 pág.
- Contreras, J. R., F. Agnolin, Y. E. Davies, I. Godoy, A. Giacchino & E. Ríos (2014). Atlas ornitogeográfico de la provincia de Formosa, República Argentina. 1º Ed. Ciudad Autónoma de Buenos Aires: Fundación de Historia Natural Félix de Azara. 450 p. (enlace)
- Coria, O. R., O. B. Quiroga, J. L. Navarro, J. Heredia, R. Torres & J. Lima (2021). Lista actualizada de las aves de Santiago del Estero, Argentina. Acta Zoológica Lilloana, 65 (1): 42-143. (enlace)
- Coria, O. R., R. Torres, M. de la Peña & J. J. Lima (2023). Aves de Santiago del Estero. Ecología y Conservación. Buenos Aires. Fundación de Hostoria Natural Félix de Azara. (enlace)
- Cuervo, P., C. Fantozzi & L. Sbriglio (2011). Registros novedosos de aves acuáticas para el noroeste y centro de la Argentina. Nótulas Faunísticas. Segunda Serie, No 63: 1-5. (enlace)
- Dabbene, R. (1918). Notas sobre una colección de aves de la Isla Martín García. Hornero, 1: 89-97. (enlace)
- Daguerre, J. B. (1922). Lista de aves coleccionadas y observadas en Rosas, F. C. S. Hornero, 2 (4): 259-271. (enlace)
- Darrieu, C. A. & A. R. Camperi (2001) Nueva lista de las aves de la provincia de Buenos Aires. Cobiobo No 3-Probiota No 2. Sec. de Política Ambiental. UNLP. Buenos Aires (enlace)
- Davis, S. E., Rocha, O. O., Sarmiento, J. & W. Hanagarth (1994). New departmental records and notes for some Bolivian birds. Bull. Brit. Orn. Club., 114 (2): 73–85. (enlace)
- de la Peña M. R. (1999). Aves Argentinas. Lista y Distribución. Ed. L.O.L.A. (Literature of Latin America). Monografía N.º 18; 244 pp. Buenos Aires. ISSN 0328-1620
- de la Peña, M. R. & R. F. Tittarelli (2011a). Guía de aves de La Pampa. Santa Rosa. Gobierno de La Pampa
- de la Peña, M. R. (2011b). Atlas ornitogeográfico de la provincia de Santa Fe. Serie Naturaleza y Conservación No 4. Pág. 1-500.
- de la Peña, M. R. (2012). Distribución y citas de aves de Entre Ríos. Serie Naturaleza y Conservación No 6. Pág. 1-206. ISBN 978-987-27043-5-3
- de la Peña, M. R. (2013). Nidos y Reproducción de las aves argentinas. Serie Naturaleza, Conservación y Sociedad No 8. Ediciones Biológica. Santa Fe, Argentina. 590 pp. ISBN 978-987-27043-7-7 (enlace)
- de la Peña, M. R. (2015). Aves Argentinas. Incluye nidos y huevos. 1º Ed. Santa Fe: Ediciones UNL; CABA: EUDEBA; Vol 1. 496 pp. ISBN 978-987-657-986-5
- de la Peña, M. R. (2025). Aves Argentinas. Descripción, Comportamiento, Reproducción y Distribución (Actualización). Tomo 1: Rheidae, Tinamidae, Anhimidae, Anatidae, Cracidae, Odontophoridae, Phasianidae. Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino”. (Nueva Serie). Pág. 1-298. ISSN 0325-3856 (enlace)
- del Hoyo, J. ed. (2020). All the Birds of the World. Lynx Edicions. Barcelona, España. 967 pp. ISBN 978-84-16728-37-4
- Di Giácomo, A. G. (2005). Aves de la Reserva El Bagual. En Di Giácomo, A. G. y S. F. Krapovickas (Eds.). Historia natural y paisaje de la Reserva El Bagual, provincia de Formosa, Argentina. Inventario de la fauna de vertebrados y de la flora vascular de un área del Chaco Húmedo. Temas de Naturaleza y Conservación, 4: 201-465. Aves Argentinas/AOP. Buenos Aires. (enlace)
- Di Giácomo, A. S., M. V. De Francesco & E. G. Coconier (editores) (2007). Áreas importantes para la conservación de las aves en Argentina. Sitios Prioritarios para la conservacion de la biodiversidad. Temas de Naturaleza y Conservación 5:1-514. CD-ROM. Edición Revisada y Corregida 1. Aves Argentinas/Asociación ornitológica del Plata, Buenos Aires. (enlace)
- Durnford, H. (1876). Ornithological Notes from the neighbourhood of Buenos Ayres. Ibis: 157-166 (enlace)
- Echevarría, A. L., N. L. Marigliano & J. M. Chani (1998). Composición y variaciones anuales de la diversidad de aves de una localidad de Bosque chaqueño serrano (Ticucho, El Cadillal, Tucumán, Argentina). Acta Zoológica Lilloana, 44 (1): 207-217 (enlace)
- Echevarría, A. L. & J. M. Chani (1999). Lista de aves del embalse El Cadillal, provincia de Tucumán. Acta Zoológica Lilloana, 45 (1): 141-145. (enlace)
- Echevarría, A. L. & J. M. Chani (2000). Estructura de las comunidades de aves acuáticas del embalse El Cadillal, Tucumán, Argentina. Acta Zoológica Lilloana, 45 (2): 219-232. (enlace)
- Echevarría, A. L. (2001). Estudios ecológicos de las aves acuáticas del embalse El Cadillal, provincia de Tucumán, Tesis doctoral, Facultad de Ciencias Naturales e Instituto M. Lillo, Universidad Nacional de Tucumán, 206, pp.
- Echevarría, A. L. & J. M. Chani (2006). Aves migratorias, la importancia del embalse El Cadillal, (Tucumán, Argentina) como sitio de tránsito e invernada. Acta Zoológica Lilloana, 50 (1-2): 97-108. (enlace)
- Echevarría, A. L., C. F. Marano, J. M. Chani & M. C. Cocimano (2008). Composición de la comunidad de aves del embalse La Angostura, Tafí del Valle, Tucumán. Acta Zoológica Lilloana, 52 (1-2): 98-105. (enlace)
- Echevarría, A. L., C. F. Marano, M. C. Cocimano, M. E. Fanjul & M. A. Cormenzana (2014). Composición y variación de la comunidad de aves del embalse El Tunal, Salta, Argentina. Acta Zoológica Lilloana, 58 (1): 80-93. (enlace)
- Echevarría, A. L., M. E. Fanjul, M. V. Martínez & A. Benavidez (2019). Avifauna de los bañados de Añatuya, área de importancia para la conservación de las aves (AICA) de Santiago del Estero, Argentina. Hornero, 34 (2): 43-49. (enlace)
- EcoRegistros (2026); Ficha: Chajá (Chauna torquata, Oken 1816); recuperado en mayo 2026 de https://www.ecoregistros.org/ficha/Chauna-torquata
- Fandiño, B. & A. A. Pautasso (2011). Avifauna del AVP La Salamandra, pp.: 62-105. En: Pautasso, A. A. 2011. La fauna y su conservación en los Bajos Submeridionales. Ediciones Biológica. Serie Naturaleza, Conservación y Sociedad No 5. Santa Fe, Argentina, 278 pp. (enlace)
- Fandiño, B., L. A. Leiva, A. A. Pautasso, H. Luna & M. Manassero (2015). Avifauna de las Reservas natural manejada “El Fisco” y privada de uso múltiple “Isleta linda”, Santa Fe, Argentina. Nuestras Aves, 60: 39-45. (enlace)
- Ferrer, D., R. Olivera, A. Elías, M. Pérez & P. Isola (2019). Lista actualizada de aves registradas en la provincia de Mendoza, Argentina. Historia Natural, Tercera Serie, 9 (2): 81-106. (enlace)
- Fiameni, M. A. (1994). Aves del Partido de Necochea. La Plata. Bs. As. Pág. 1-39.
- Fiora, A. i(1939) Distribución geográfica de las especies de aves capturadas en la provincia de Jujuy y zonas limítrofes. Physis, 16: 271-289.
- Filipello, A. M. & J. López de Casenave (1993). Variación estacional de la comunidad de aves acuáticas de la Reserva Costanera Sur. Rev. Mus. Arg. Cienc. Nat. B. Rivadavia. Ecología, 4 (3): 1-15. (enlace)
- Fiora, A. (1939) Distribución geográfica de las especies de aves capturadas en la provincia de Jujuy y zonas limítrofes. Physis, 16: 271-289.
- Fontana, C. S., C. V. Cademartori, R. A. Ramos, C. J. Drehmer & A. E. Tavares (1994). Abundância relativa de Chauna torquata (Oken,1816) (Aves, Anhimidae) em terras úmidas do Rio Grande do Sul, Brasil. Biociências, 2: 125-133.
- Fraga, R. M. (2001). The avifauna of Estancia San Juan Poriahú, Iberá marshes, Argentin: check-list and some natural history notes. Cotinga, 16: 81-86. (enlace)
- Gasparri, B., D. Del Río, A. Earnshaw, C. Henschke, G. Bryant, L. Cristaldo, A. Faccioli, J. M. Gentiles, I. Galiano Guirao, M. Vitale, G. Rodríguez Tourón & G. Suárez (2018). Lista comentada de las aves del Parque Natural Municipal Ribera Norte, San Isidro, provincia de Buenos Aires, Argentina. Nótulas Faunísticas, 234: 1-43. (enlace)
- Gavio, H. S. (1944). Una excursión a los cañadones de la estancia Cari-Lauquén en Santo Domingo. Hornero, 8: 494-511 (enlace)
- Gelain, M. A. & J. Tolosa (2011a). Aves de la provincia de Río Negro. Lista sistemática. Xolmis CRO, 6: 1-37
- Gelain, M. A. & R. Pereyra Lobos (2011b). Lista de aves de la provincia de San Juan, Argentina. Xolmis CRO, 3: 1- 12.
- Giacomelli, E. (1907). Catálogo sistemático de la avifauna riojana. Anales de la Sociedad Científica Argentina, (1) 63: 280-301. (enlace)
- Giacomelli, E. (1923). Catálogo sistemático de las aves útiles y nocivas de la provincia de La Rioja. El Hornero, 3 (1): 66-84. (enlace)
- Giai, A. G. (1950). Notas de viajes. Por el norte occidental de Santa Fe y por el norte de Misiones. Hornero, 9 (2): 121-164. (enlace)
- Gibson, E. (1880). Ornithological notes from the neighbouhood of Cape San Antonio, province of Buenos Ayres. Ibis (4th ser.) 4: 1-38. (enlace)
- Gibson, E. (1920). Further ornithological notes from the neighburhood of Cape San Antonio, Province of Buenos Ayres. Part III. Phoenicopteridae-Rheidae. Ibis Serie XI, , Vol. II: 1-97. (enlace)
- Giraudo, A. R. , A. Bortoluzzi & V. Arzamendia (2006). Vertebrados tetrápodos de la Reserva y Sitio Ramsar “Esteros del Iberá” (Corrientes, Argentina): análisis de su composición y nuevos registros para especies amenazadas. Natura Neotropicalis, 37 (1-2): 1-20. (enlace)
- González, J. (2018). Chajá (Chauna torquata) en Malargüe, Mendoza. Nuestras Aves, 63, 20-21. (enlace)
- Gore, M. E. J. & A. R. M. Gepp (1978). Las Aves del Uruguay. Mosca Hermanos S.A., Montevideo, Uruguay.
- Grant, C. H. B. (1911). List of Birds collected in Argentina, Paraguay, Bolivia y Southern Brasil, with field notes. Ibis, 1911: 317-350. (enlace)
- Gyldenstolpe, N. (1945). A Contribution to the Ornithology of Northern Bolivia. Kungl. Svensk. Vetensk. Akad. Handl., (3) 23 (1): 1-300.
- Haedo Rossi, J. A. (1970). Notas ornitológicas IX. Observaciones sobre el “Chajá”, “Ñandú petiso”, “Ñandú gris” y “ Ñandú blanco” en cautividad. Acta Zool. Lilloana, 26 (3): 25-32.
- Hartert, E. & S. Venturi (1909). Notes sur les oiseaux de la République Argentine. Novit. Zool., 16: 159-267. (enlace)
- Heredia, J. (2011). Informe ornitológico sobre las Lagunas Saladas de Santiago del Estero, Argentina. Dirección General de Bosques y Fauna. Santiago del Estero.
- Holland, A. H. (1890). On some birds of the Argentine Republic. Wiht Notes by P. L. Sclater. Ibis, 1890: 424-428 (enlace)
- Holland, A. H. (1892). Short notes on the birds of Estancia Espartillar, Argentina Republic. Ibis, 1892: 193-214. (enlace)
- Isacch, J. P. & M. M. Martínez (2001). Estacionalidad y relaciones con la estructura del hábitat de la comunidad de aves de pastizales de Paja colorada (Paspalum quadrifarium) manejados con fuego en la provincia de Buenos Aires, Argentina. Ornitología Neotropical, 12: 345-354. (enlace)
- Josens, M. L., Pretelli, M. G. & A. H. Escalante (2009). Censos de aves acuáticas en sus colonias reproductivas en lagunas del sudeste de la provincia de Buenos Aires. Hornero, 24 (1): 7–12. (enlace)
- Juárez, M. (2021). Variación estacional de la riqueza y frecuencia relativa de aves acuáticas de la laguna “La Picasa”, Santa Fe, Argentina. Nuestras Aves, 66: 34-40. (enlace)
- Klimaitis, J. F. (1975). Lista sistemática de aves del Partido de Berisso (Bs. As.). Parte I: No passeriformes. Hornero, 11: 271-280. (enlace)
- Klimaitis, J. F. & F. N. Moschione (1987). Aves de la Reserva Integral de Selva Marginal de Punta Lara y sus Alrededores. Dirección de Servicios Generales del Ministerio de Economía de la Provincia de Buenos Aires, Argentina.
- Lane, D. F. (2014). New and noteworthy records of birds in Bolivia. Cotinga, 36:56–67.
- Laubmann, A. (1930). Vögel en Wissenschaftliche Ergebnisse der Deutschen Gran Chaco-Expedition. Verlag von Strecker und Schröder, Stuttgart.
- Lepage, D. (2003). Avibase-La base de datos World Bird. Ficha: Chajá común (Chauna torquata, Oken, 1816); recuperado en mayo 2026 de https://avibase.bsc-eoc.org/species.jsp?avibaseid=91194BE324AB9F4C
- López-Lanús, B. (1997). Inventario de las aves del Parque Nacional «Río Pilcomayo» (Formosa, Argentina). Monografía especial Nº 4; 76 pp. Ed. L.O.L.A. (Literature of Latin America)., Buenos Aires, Argentina. ISSN: 0328-1620
- López-Lanús, B., Grilli, P., Coconier, E., Di Giacomo, A. & R. Banchs (2008). Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas/AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina, 64.
- López-Lanús, B., A. Galimberti, C. Giarduz, H. Luna, M. Manassero, A. A. Pautasso, M. P. Ducommun, A. A. G. Saigo, P. Petracci, G. A. Marteleur & M. R. de la Peña (2013a). Inventario focal de fauna de las estancias El Estero, La Norma, El Matrero y La Elena en el sitio piloto San Javier: Santa Fe, Argentina. En pp. 135-169/277-306/331-347/507,511-513: G. D. Marino, F. Miñarro, M. E. Zaccagnini y B. López-Lanús (eds.). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. (enlace)
- López-Lanús, B., A. S. Di Giacomo, A. Azpiroz, P. Haynes, A. Galimberti, A. Keyel, A. Ocampo, R. Güller, R. Moller Jensen, M. Mattalia, H. Cardoso, C. Giarduz, G. Papini y A. G. Di Giacomo (2013b). Inventario focal de fauna de las estancias La Higuera, María Concepción, La Serena y Virocay en el sitio piloto Aguapey: Corrientes, Argentina. En pp. 179-223/277-293/307-319/331-347/490,494-506: G. D. Marino, F. Miñarro, M. E. Zaccagnini y B. López-Lanús (eds.). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. (enlace)
- López-Lanús, B., P. Grilli, E. Carini, M. Montero & J. Casillo (2013c). Inventario focal de fauna de las estancias La Catalina, La Aurora del Palmar, La Filiberta y Campo Bajo en el sitio piloto Gualeguaychú: Entre Ríos, Argentina. En pp. 231-252/277-293/321-347/525-538: G.D. Marino, F. Miñarro, M.E. Zaccagnini y B. López-Lanús (eds.). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. (enlace)
- López Lanús, B. & P. Mencia (2017). Diversidad, abundancia y estado de conservación de las aves de los riachos Monte Lindo y Pilagá en el este de la provincia de Formosa, Argentina. Nótulas Faunísticas (segunda serie), 228: 1-29. (enlace)
- Lozano, J. & A. F. Malo (2013). Relationships between human activity and richness and abundance of some bird species in the Paraguay River (Pantanal, Brazil). Ardeola, 60 (1): 99–112.
- Lucero, M. M. (1983). Lista y distribución de aves y mamíferos de la provincia de Tucumán. Miscelánea, Nº 75. Fund. M. Lillo. Tucumán. 61 pp. (enlace)
- Lucero, F. (2012). Nuevos registros y distribución de aves par alas provincias de San Juan, Mendoza y La Rioja. Parte II. Nótulas Faunísticas, Segunda Serie, 97: 1-10. (enlace)
- Lucero, F. (2013a). Listado de las aves observadas en la Reserva Natural Municipal Articulada: Laguna Guanacache, Laguna del Toro y Bañados del Carau, provincia de San Juan y Laguna Seca y los Bañados del Tulumaya, provincia de Mendoza, Argentina. Nótulas Faunísticas (segunda serie), 137: 1-16. (enlace)
- Lucero, F. (2013b). Aves de Sarmiento, provincia de San Juan. Edición del autor. 1a Ed., Cienaguita, 334 pp.
- Lucero, F. & F. Izasa (2015). Primeros registros, nuevas localidades, observación de aves con documentación escasa y evidencias de nidificación para las provincias de Mendoza, San Juan, La Rioja, Catamarca y Tucumán, Argentina. Ecoregistros Revista, 5 (8): 18-48. (enlace)
- Lucero, F. (2020a). Nidificación del Chajá (Chauna torquata) en Guanacache (Sirio Ramsar), Provincia de Mendoza, presencia en la región de Cuyo, pérdida de hábitat y sitios dispersos de ocupación de una especie en peligro. Ecoregistros Revista, 10 (6): 26-32. (enlace)
- Lucero F. H. (2020b). Distribución y Nidificación de las aves en la provincia de San Juan, Argentina. NO PASSERIFORMES II. (del Chajá a las gallaretas). (enlace)
- Luna, N., M. Libua, M. O. Gallegos, M. Cueva & L. E. Rodríguez (2008). Redescubrimiento del Ganso de monte Neochen jubata (Spix, 1825) para la avifauna argentina. Nótulas Faunísticas, 24: 1-13. (enlace)
- Mancini, J. A. (2011). Inventario de las aves del Parque Escolar Rural Enrique Berduc. Entre Ríos.
- Marateo, G., H. Povedano & J. Alonso (2009). Inventario de las aves del Parque Nacional El Palmar, Argentina. Cotinga, 31: 47-60 (enlace)
- Marelli, C. A. (1918). Aves de Curuzú Cuatiá. (Corrientes). Hornero, 1: 74-80. (enlace)
- Marino, G.D., F. Miñarro, M.E. Zaccagnini & B. López-Lanús (eds.). (2013). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. (enlace)
- Martínez, F., F. Lucero, R. Calí, D. Valdés, D. Ferrer & J. C. Chébez (2009). Registros novedosos de aves para las provincias de Mendoza y San Juan. Nótulas Faunísticas. Segunda serie, Nº 35: 1-9. (enlace)
- Mérida, E. & A. Bodrati (2006). Consideraciones sobre la conservación de los talares de barrancas del nordeste de Buenos Aires y descripción de las características de un relicto en Baradero. Pp.: 71-82, en : Mérida, E. y J.Athor (Editores). Talares bonaerenses y su conservación. Fundación de Historia Natural “Félix de Azara”. Buenos Aires. ISBN-13: 978-987-22121-5-5 (enlace)
- Morici, A. (2012). Las aves del área Natural Protegida “Dique Roggero” (Buenos Aires, Argentina). Biológica, 15: 24-36 (enlace)
- Narosky, T. & A. G. Di Giacomo (1993). Las aves de la provincia de Buenos Aires, distribución y estatus. Asoc. Orn. del Plata. Vazquez Mazzini Editores, L.O.L.A., Buenos Aires
- Narosky, T. & A. Martelli (1995). Una nueva visita al este de Formosa. Nuestras Aves, 31: 28-29 (enlace)
- Narosky, T. & D. Yzurieta (2010). Birds of Argentina and Uruguay: a field guide. Vázquez Mazzini Editores, Buenos Aires, Argentina. ISBN 950-99063-6-0
- Nellar Ramonella, M. M. (1993). Aves de la provincia de San Luis. Lista y distribución. Ed. Mus. Priv. Cienc. Nat. E Inv. Ornit. “G. E. Hudson”. San Luis. Págs. 98. ISBN: 9789504350491
- Olmos, F., L. F. Silveira & G. A. Benedicto (2011). A contribution to the ornithology of Rondônia, southwest of the Brazilian Amazon. Revista Brasileira de Ornitologia, 19 (2): 200–229. (enlace)
- Nores, M. & D. Yzurieta (1980). Aves de ambientes acuáticos de Córdoba y centro de Argentina. Sec. de Estado de Agricultura y Ganadería. Acad. Nac. Cienc. de Córdoba. 235 pp.
- Nores, M. (1996). Avifauna de la provincia de Córdoba. Fauna, 1: 255-337.
- Ochoa de Masramón, D. (1983). Lista de aves del nordeste de San Luis. Hornero, 12 (1extra): 77-87. (enlace)
- Olrog, C. Ch. (1959). Las aves argentinas. Una guía de campo. Universidad Nacional de Tucumán. Instituto «Miguel Lillo». Tucumán, Argentina.
- Ordano, M., A. Bortoluzzi, M. Chatellenaz & L. Biancucci (2008). Respuesta a corto plazo de las aves a la quema y al pastoreo en pastizales del Parque Nacional Mburucuyá, Argentina. INSUGEO, Miscelánea, 17 (2): 425-446. (enlace)
- Ortiz, S. G., H. A. Gómez, R. V. Blanco Fager,, L. Videla & E. Godoy Luna (2016). Aves de las diferentes ecorregiones, San Juan, Argentina. En: Martínez Carretero E. (ed.). San Juan Ambiental. 494 pp.
- Pagano, L. G. & E. Mérida (2009). Aves del Parque Costero del Sur. Pp. 200-244 en: Athor J (ed) Parque Costero del Sur. Naturaleza, conservación y patrimonio cultural. Fundación Félix de Azara, Buenos Aires, Argentina. (enlace)
- Pagano, L. G., E. A. Jordan, J. I. Areta, R. F. Jensen & I. Roesler (2012). Aves de la Reserva Natural Punta Lara. En 97-143: Roesler, I. y M.G. Agostini (eds). Inventario de los Vertebrados de la Reserva Natural Punta Lara, provincia de Buenos Aires, Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 8. Buenos Aires, Argentina. (enlace)
- Parker, T. A. (1982). Observations of some unusual rainforest and marsh birds in southeastern Peru. Wilson Bull., 94 (4): 477–493. (enlace)
- Pautasso, A. A. (2002). Aves de la Reserva Urbana de la Ciudad Universitaria UNL “El Pozo”, Santa Fe. Argentina. Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino”, 8: 1-12. (enlace)
- Pearson, D. L. (1975). Range Extension and New Records for Bird Species in Ecuador, Perú and Bolivia. Cóndor, 77 (1): 96-99. (enlace)
- Peltzer, P. M. (1998). Inventario de flora y fauna del Área Natural Protegida: Islote municipal del valle aluvial del Paraná. Natura Neotropicalis, 29: 74-78 (enlace)
- Pereyra, J. A. (1923) Las aves de la región ribereña de la provincia de Buenos Aires. Hornero, 3 (02) : 159-174 (enlace)
- Pereyra, J. A. (1927). Segunda lista de aves colectadas en la región ribereña de Buenos Aires. Hornero, 4: 23-34 (enlace)
- Pereyra, J. A. (1938). Aves de la zona ribereña nordeste de la provincia de Buenos Aires. Memorias del Jardín Zoológico. La Plata. Tomo 9, parte 2: 1-304. (enlace)
- Pereyra, C. B. de. (1940). Nota sobre el Chajá. Chauna torquata (Swainson). Hornero, 7: 378-381. (enlace)
- Planqué, R., Vellinga, W. P., Pieterse, S., Jongsma J. & R. de By (Xeno-canto Foundation). Ficha: Southern Screamer · Chauna torquata · (Oken, 1816); recuperado en mayo 2026 de https://xeno-canto.org/species/chauna-torquata
- Pinto, O. M. O. (1964). Ornitologia Brasiliense. Volume 1. Departamento de Zoologia da Secretaria da Agricultura do Estado de São Paulo, São Paulo, Brazil.
- Pugnali, G. & P. Chamorro (2008). Lista de aves de la Reserva Costanera Sur. Aves Argentinas. Buenos Aires. (enlace)
- Pujalte, J. C., A. R. Reca, A. Balabusic, P. Canevari, L. Cusato & V. P. Fleming (1995). Unidades ecológicas del Parque Nacional Río Pilcomayo. Anales de Parques Nacionales, 16: 1-185. (enlace)
- Quiroga, M. A & H. R. Hadad (2004). Diversidad de aves y macrófitas en diferentes comunidades vegetales de un humedal. Revista FABICIB, (8): 435-57. (enlace)
- Rand, A. L. (1954); On the spurs on Bird’s wings. Wilson Bulletin, Vol. 66: Iss. 2 , Article 8. (enlace)
- Reed, C. S. (1921). Las aves de caza de la provincia de Mendoza. Revista Chilena de Historia Natural, 25: 203-220.
- Renard, A. (1920). Notas sobre aves de Cañuelas (Prov. de Buenos Aires). Hornero, 2: 58-60. (enlace)
- Rodríguez Graves, V., A. Pautasso, P. Preliasco, M. V. Del Sastre & F. Miñarro (2009). Relevamiento ambiental de la Estancia Isleta Linda. Provincia de Santa Fe. Fundación Vida Silvestre Argentina. (enlace)
- Roig, V. G. (1965). Elenco sistemático de los mamíferos y aves de la provincia de Mendoza y notas sobre su distribución geográfica. Boletín de Estudios geográficos, (49). (enlace)
- Rumboll, M. A. E. (1975). Espolones metacarpales del Chajá (Chauna torquata). Hornero, 11: 316-317.
- Ruschi, A. (1979). Aves do Brasil. Volume 1. Editora Rios, São Paulo, Brazil.
- Salvador, S. A. (1983). La avifauna en Villa María y sus alrededores. Escuela Normal “ Víctor Mercante” . Inst. Sec. “Bernardino Rivadavia”. Villa María, Córdoba. Argentina.
- Salvador, S. A., L. G. Pagano & F. G. Di Sallo (2016). Primeros registros provinciales de reproducción de algunas aves en Argentina (Catamarca y La Rioja). Nuestras Aves, 61: 32-35. (enlace)
- Salvador, S. A., L. A. Salvador & C. Ferrari (2017). Aves de la provincia de Córdoba. Distribución e Historia Natural. Ed. DP. Argentina. S.A.
- Sanzín, R. (1918). Lista de aves mendocinas. Hornero, 1: 147-152. (enlace)
- Saporiti, E. J. (1947). Biología del Chajá, Chauna torquata, Oken. Anales Sociedad Científica Argentina, 144: 434-445. (enlace)
- Schulenberg, T. S., D. F. Stotz, D. F. Lane, J. P. O’Neill & T. A. Parker (2007). Birds of Peru. Princeton University Press, Princeton, NJ, USA. ISBN: 13: 978-0-691-04915-1
- Scott, D. A. & M. Carbonell, Editors (1986). A Directory of Neotropical Wetlands. International Wetland Research Bureau, Slimbridge and International Union for the Conservation of Nature, Cambridge, UK.
- Segura, L. N., N. Bogado, C. A. Darrieu & D. Montalti (2014). La colección de huevos “Pablo Girard” del Museo de La Plata. Probiótica. FCN y M, UNLP, La Plata, Argentina, Serie Técnica y Didáctica, 27: 1-14. (enlace)
- Serié, P. & C. H. Smyth (1923). Notas sobre aves de Santa Elena (E. Ríos). Hornero, 3: 37-55. (enlace)
- Servat, G. P. & D. L. Pearson (1991). Natural history notes and records for seven poorly known bird species from Amazonian Peru. Bull. Brit. Orn. Club, 111 (2): 92–95. (enlace)
- S.I.B. (Sistema de Información de Biodiversidad). Administración de Parques Nacionales. Ficha: Chajá (Chauna torquata); Recuperado en junio 2026 de https://sib.gob.ar/especies/chauna-torquata
- Sick, H. (1993). Birds in Brazil. A natural history. Princeton University Press, Princeton, New Jersey. ISBN: 0-691-08569-2. (enlace)
- Sick, H. (1997). Ornitologia Brasileira. Editora Nova Fronteira, Rio de Janeiro, Brazil. 912 pp. ISBN 85-209-0816-0
- Siegenthaler, G. B. (Coord.) (2004). Relevamiento de los vertebrados de la provincia de La Pampa. Inventario integrado de los recursos naturales de la provincia de La Pampa. Clima, geomorfología, suelo, vegetación y fauna de vertebrados. Reedición. INTA.UNLP. Gobierno de La Pampa, CD, multimedia.
- Smyth, C. H. (1927). Descripción de una colección de huevos de aves argentinas. Hornero, 4 (1): 1-16. (enlace)
- Soothill, E. & P. Whitehead (1978). Wildfowl of the World. Peerage Books, London. (enlace)
- Stenberg, C. (1869). Notizen aus der Vogelwelt von Buenos Ayres. Journ. Für Ornith., 1869: 174-193, 257-278 (enlace)
- Stonor, C. R. (1939). Notes on the breeding habits of the Common Screamer (Chauna torquata). Ibis, 81:45–49.
- Tajer, L., F. Rizzo & E. Palombarini (2007). Relevamiento de las aves del Parque Plaza Montero, Reserva Natural Urbana de Las Flores, Provincia de Buenos Aires, Argentina. Aves Argentinas/AOP. Pp: 1-19. (enlace)
- Torres, R. & P. Michelutti (2006). Aves acuáticas. Banados del Rıo Dulce y Laguna Mar Chiquita (Córdoba, Argentina) (EH Bucher). Academia Nacional de Ciencias, Córdoba, Argentina, 237-249. (enlace)
- Veselovsky, Z. (1986). Beitrag zur Kenntnis des Brutverhaltens des Schopfwehrvogels (Chauna torquata). Zool. Garten., 56 (4-5): 363-384.
- Weller, M. W. (1967). Notes on some marsh birds of Cape San Antonio, Argentina. Ibis, 109 (3): 391–411.
- Weller, M. (1999). Wetland birds: habitat resources and conservation implications. Cambridge University Press, New York, New York. (enlace)
- Wetmore, A. (1926). Observations on the birds of Argentina, Paraguay, Uruguay and Chile. United States National Museum Bulletin, 133:1–448. (enlace)
- White, E. W. (1882). Notes on birds collected in the Argentine Republic. Proc. Zool. Soc. London, 591-629. (enlace)
- Wilson, A. S. (1926). Lista de aves del sur de Santa Fe. Hornero, 3: 349-362 (enlace)
- Yzurieta, D. (1995). Manual de reconocimiento y evaluación ecológica de las aves de Córdoba. Ministerio de Agricultura, Ganadería y Recursos Renovables. Córdoba.
- Zapata, A. R. P. (1975). Aves observadas en la proximidad de la confluencia de los ríos Uruguay y Gualeguaychú, provincia de Entre Ríos. Hornero, 11: 291-304 (enlace)
- Zotta, A. R. (1932). Notas sobre el contenido estomacal de algunas aves. Hornero, 5: 77-81. (enlace)
- Zuberbühler, E. A. (1971). Observaciones sobre las aves de la provincia de Buenos Aires. Hornero, 11: 98-112. (enlace)
-
Chajá | Conservación
ESTADO DE CONSERVACIÓN: PREOCUPACIÓN MENORLa Chauna torquata (Chajá) es considerado una especie de “Preocupación Menor” por BirdLife International debido a su amplia distribución geográfica, gran tamaño poblacional y tendencia poblacional estable, factores que lo mantienen lejos de los umbrales de amenaza establecidos (BirdLife International 2010). En este sentido, no cumple criterios de vulnerabilidad asociados a reducciones significativas de distribución, población o declives superiores al 30 % en el corto o mediano plazo, lo que refuerza su clasificación como especie no amenazada (Brady, 2020).
Aunque no se encuentra en riesgo, esta especie presenta interacciones relevantes con la actividad humana: puede ser domesticada y es utilizada como animal de guardia debido a su potente vocalización, mientras que en contextos agrícolas es frecuentemente considerada una especie problemática por su comportamiento gregario. Asimismo, la desecación de humedales afecta sus áreas de reproducción, aunque la especie muestra cierta capacidad de adaptación a estas modificaciones ambientales (Carboneras, 1992:533).
En relación con sus depredadores, se han registrado diferentes amenazas según las etapas del ciclo de vida. En estado adulto, se ha documentado la depredación por el zorro pampa o gris en Maipú, Buenos Aires (Argerich, E., 31/12/2017; EcoRegistros 2019). En etapas tempranas, los huevos y pichones pueden ser depredados por la musurana en Formosa (Gallardo et al., 2006) por el curiyú en Formosa (Waller et al., 2007).En Argentina, a pesar de no estar considerada una especie amenazada (López Lanús et al., 2008:46), enfrenta múltiples presiones antrópicas que pueden generar consecuencias significativas sobre sus poblaciones. Entre ellas se destaca la caza sostenida, que ha provocado disminuciones locales evidentes en áreas de persecución activa y un descenso general en regiones como la Provincia del Chaco (Contreras et al., 1990). Además, su sensibilidad a las modificaciones drásticas del hábitat, particularmente aquellas vinculadas a la alteración de recursos hídricos, constituye un factor crítico. También se señala su exposición a los efectos directos e indirectos de agroquímicos debido a su frecuente presencia en arrozales, lo que incrementa su vulnerabilidad ambiental y refuerza la necesidad de medidas de protección legal (Contreras et al., 2014:74).
Finalmente, las estimaciones poblacionales globales para la Argentina indican un rango aproximado entre 100.000 y 1.000.000 de individuos (Delaney & Scott, 2002; Di Giácomo et al., 2007:496), lo que confirma su amplia abundancia relativa a escala regional.Estado de conservación en Argentina (SIB)
No Amenazada (NA): según AA y SAyDS, 2008 (Aves Argentinas y Secretaría de Ambiente y Desarrollo Sustentable). López-Lanús, B., P. Grilli, E. Coconier, A. Di Giacomo y R. Banchs. Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas /AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina. 1 ed. AOP: Buenos Aires, Argentina
No Amenazada (NA): según MADS y AA, 2017 (Ministerio de Ambiente y Desarrollo Sustentable de la Nación y Aves Argentinas). Categorización de las Aves de la Argentina según su estado de conservación. Informe del Ministerio de Ambiente y Desarrollo Sustentable de la Nación y de Aves Argentinas. 146 p. 2017
Preocupación Menor (LC): según UICN (API) (Unión Internacional para la Conservación de la Naturaleza). Lista Roja de la UICN publicada en https://www.iucnredlist.org. Actualización en tiempo real -
Chajá | Distribución
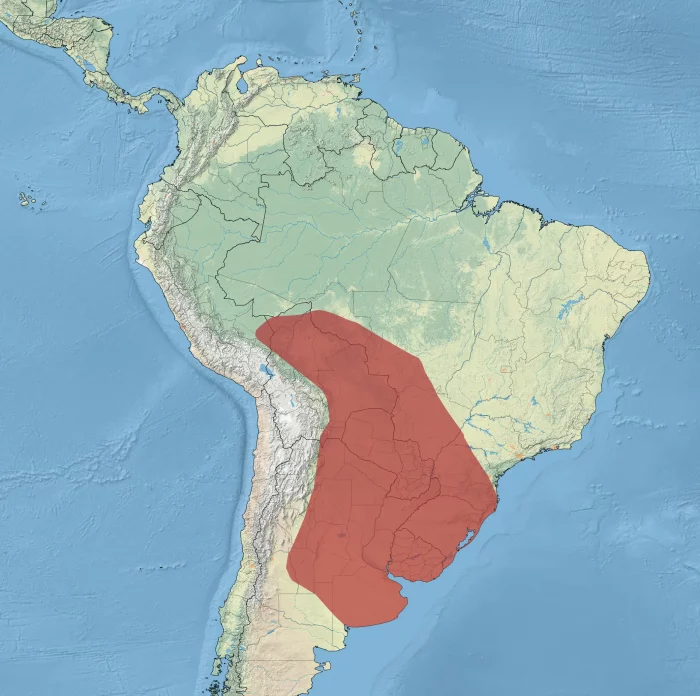
© Animalia. Algunos derechos reservados Es una especie endémica de Sudamérica (Contreras, 2014:75), cuya distribución se extiende principalmente por la Cuenca del Plata y regiones adyacentes, con registros aislados en la Amazonia peruana (Servat & Pearson, 1991:92; Contreras, 2014:75). Su rango de distribución comprende Bolivia, Paraguay, Uruguay, Argentina y Brasil (Sick, 1997:242; Olrog, 1979:46).
En Bolivia, la especie se distribuye ampliamente en el norte y este de los departamentos de Beni, Santa Cruz y Tarija (Blake, 1977:211), con presencia general desde el país hacia el sur hasta el norte de Argentina y hacia el este hasta los estados brasileños de Mato Grosso y Rio Grande do Sul (Davis et al., 1994:76; Brumfield et al., 2004:385). Asimismo, se ha registrado en Paraguay y Uruguay (Gore y Gepp, 1978), así como en el sur de Brasil, desde Mato Grosso hasta el oeste de São Paulo y Rio Grande do Sul (Blake, 1977:211; Sick, 1997:242). En Argentina, su distribución alcanza desde las regiones septentrionales hasta las provincias de Mendoza, La Pampa y Buenos Aires (Blake, 1977:211; Olrog, 1979:46; Josens et al., 2009:9).
Existen además registros erráticos en el sudeste de Perú (Parker, 1982:478; Servat & Pearson, 1991:92), lo que ha llevado a señalar su presencia ocasional en la Amazonia peruana (Contreras, 2014:75).
La especie habita preferentemente ambientes palustres, marismas y lagunas con abundante vegetación acuática. En Rio Grande do Sul se la considera un residente común en humedales y lagos con vegetación emergente, particularmente en las zonas litorales y en sectores occidentales de la depresión central del estado (Belton, 1984:477).
Aunque generalmente se considera una especie no migratoria, estudios terrestres y aéreos realizados en Rio Grande do Sul evidenciaron fluctuaciones estacionales en la abundancia. Las poblaciones costeras disminuyen entre julio y noviembre, coincidiendo con incrementos en localidades del interior, lo que sugiere la existencia de movimientos locales o desplazamientos regionales (Fontana et al., 1994).HÁBITATLa especie se encuentra estrechamente asociada a ambientes de humedales tanto tropicales como subtropicales, incluyendo lagos, marismas, lagunas, esteros y pantanos, generalmente en áreas con vegetación acuática y, en algunos casos, con presencia de árboles dispersos (Brady, 2020; Olrog, 1979:46).
Habita una amplia variedad de humedales continentales, tales como lagunas, esteros, bañados y sabanas húmedas, así como bosques abiertos e inundados y arrozales (Contreras et al., 2014:74; de la Peña, 2025:86). Asimismo, puede utilizar ambientes antropizados o modificados, como campos inundados y áreas agrícolas.
A escala regional, se ha documentado su presencia en lagunas, bañados, campos y diques en la provincia de Córdoba, con excepción de la región altoserrana, donde no se registra. Las mayores densidades poblacionales se concentran en las zonas este, centro y sur de dicha provincia (Salvador et al., 2017:16).
En Argentina, también ocupa una diversidad de microhábitats acuáticos como represas, orillas de caminos, bañados, esteros y sectores con aguas abiertas en riachos, incluyendo pozas y claros (Di Giácomo, 2005:230). En conjunto, estos registros confirman su fuerte dependencia de sistemas de humedales y su amplia tolerancia a distintos tipos de cuerpos de agua someros (Olrog, 1979:46; Contreras et al., 2014:74).
Ecorregiones: Pampa, Chaco y Delta e Islas del Paraná , Espinal Esteros del Iberá y Yungas (Barnett & Pearman, 2001:30)DISTRIBUCIÓN GEOGRÁFICA EN ARGENTINAEn Argentina se distribuye desde el norte del país hasta el sureste de San Juan y abarcando también el este de Mendoza, La Pampa y Río Negro (de la Peña, 2025:91). En términos generales, ocupa gran parte del territorio argentino no serrano, alcanzando las provincias de Buenos Aires, La Pampa, Córdoba, San Luis, Mendoza y Río Negro (Contreras, 2014:75).
Son residentes durante todo el año. En la Reserva El Bagual (Formosa), la especie ha sido considerada residente permanente, con registros correspondientes a todos los meses del año (Di Giácomo, 2005:230). Los antecedentes para la provincia de Formosa incluyen las observaciones realizadas por Wetmore (1926) entre el 10 y el 21 de agosto de 1920 en el Riacho Pilagá, donde se colectó un macho adulto el 16 de agosto, así como un ejemplar obtenido por la expedición Krieg en Lapango durante septiembre de 1925 (Laubmann, 1930). Posteriormente, Narosky y Martelli (1995) la señalaron como una especie común en la estancia Clarín durante octubre de 1991. Su presencia también ha sido documentada en el Parque Nacional Río Pilcomayo (Pujalte et al., 1995; López Lanús, 1997) y en la Reserva Natural Formosa (Chebez et al., 1998).
De acuerdo con registros más recientes, la especie se encuentra presente en diecinueve provincias: Buenos Aires, Catamarca, Chaco, Córdoba, Corrientes, Entre Ríos, Formosa, Jujuy, La Pampa, La Rioja, Mendoza, Misiones, Río Negro, Salta, San Juan, San Luis, Santa Fe, Santiago del Estero y Tucumán (Lucero, 2020:26; de la Peña, 2019). Dentro de este amplio rango de distribución, se ha documentado su nidificación en al menos doce provincias, incluyendo Jujuy, Salta, Formosa, Chaco, Santiago del Estero, Corrientes, Santa Fe, Córdoba, Entre Ríos, San Luis, Buenos Aires y La Pampa (Salvador, 2017).Citas y observaciones en las provincias
-
Chajá | Comportamiento
El Chajá (Chauna torquata) es una especie de hábitos predominantemente terrestres y diurnos. Aunque posee capacidad para nadar, generalmente prefiere desplazarse caminando, comportamiento que ha llevado a describirlo como un ave terrícola y caminadora (Brady, 2020; Di Giacomo, 2005:230). Su desplazamiento terrestre se caracteriza por movimientos lentos y un andar majestuoso, acompañado de una postura erguida, una actitud vigilante y una apariencia de continuo estado de alerta (de la Peña, 2025:86; Saporiti, 1947:434). Belton (1984:479) señala que los individuos suelen mantener el cuello vertical, rasgo que refuerza esta impresión de vigilancia permanente.
Se observa con mayor frecuencia en parejas, condición que diversos autores consideran característica de su comportamiento habitual (Belton, 1984:479; Contreras et al., 2014:74; Salvador et al., 2017:16). Saporiti (1947:434) destaca incluso una marcada tendencia a la vida en pareja, señalando que los individuos suelen mantener una existencia independiente y monógama. No obstante, fuera de la temporada reproductiva pueden reunirse en grupos de tamaño variable y formar grandes concentraciones. Carboneras (1992:530) menciona que durante este período la especie se vuelve social, alimentándose en bandadas de hasta 100 individuos y descansando en grupos. Asimismo, Belton (1984:479) registró una concentración de 181 ejemplares en unas pocas hectáreas, mientras que Di Giacomo (2005:230) señala agrupaciones dispersas que superan el centenar de individuos durante el otoño o después de intensas lluvias en bañados y grandes esteros. Sick (1997:242) también observó grupos de individuos aparentemente no reproductivos que deambulan pacíficamente en distintas épocas del año. En contraste, Salvador et al. (2017:16) indican que los grupos de más de dos individuos son poco frecuentes, generalmente integrados por cuatro a ocho ejemplares.
La especie frecuenta ambientes abiertos asociados a humedales. Habitualmente se la observa vadeando en marismas poco profundas o descansando en charcas y pantanos de escasa profundidad (Belton, 1984:479; Carboneras, 1992:530; Sick, 1997:242). También utiliza con frecuencia árboles, arbustos y otras estructuras elevadas como sitios de reposo, especialmente para dormir o vigilar el entorno (Belton, 1984:479; de la Peña, 2025:86; Di Giacomo, 2005:230; Sick, 1997:242). Cuando se siente amenazada, suele refugiarse en las copas de los árboles, desde donde emite fuertes vocalizaciones de alarma (Brady, 2020).
El vuelo constituye una de las capacidades más notables de la especie. Aunque el despegue puede parecer dificultoso debido a su tamaño corporal, una vez en el aire demuestra gran habilidad para el planeo (de la Peña, 2025:86; Brady, 2020). Durante el ascenso realiza vigorosos aleteos que producen un ruido característico, alcanzando posteriormente alturas considerables gracias al aprovechamiento de corrientes térmicas (Belton, 1984:479; de la Peña, 2025:86). Gibson (1880:166) describió su vuelo como lento y compuesto por amplios y potentes movimientos alares. Di Giacomo (2005:230) observó que frecuentemente comparte las corrientes térmicas con otras aves de gran porte, como los Jabirúes (Jabiru mycteria), Tuyuyúes (Mycteria americana) y Jotes Cabeza Negra (Coragyps atratus). Sin embargo, Sick (1997:242) señala que durante el vuelo puede maniobrar torpemente, manteniendo las patas parcialmente colgantes.
Finalmente, se ha documentado un caso de depredación sobre la especie: en el estómago de una musurana (Boiruna maculata) colectada en la REB y depositada en la colección de la FML se halló un pichón de Chajá de aproximadamente una semana de vida (Gallardo et al., en prensa), registro citado por Di Giacomo (2005:230).
VOCALIZACIONESLas vocalizaciones del Chajá (Chauna torquata) constituyen uno de los rasgos más distintivos de la especie. Su voz es extremadamente potente, de gran alcance y fácilmente audible a largas distancias; de hecho, se ha señalado que sus llamados, semejantes a los de los gansos, pueden escucharse a más de tres kilómetros (Sick, 1997:242; Belton, 1984:477). Asimismo, las vocalizaciones han sido descritas como fuertes, poco melodiosas y de carácter trompeteante (Brady, 2020). Los individuos emiten llamados intensos cuando son perturbados y pueden vocalizar incluso durante la noche (Belton, 1984:479; Di Giacomo, 2005:230). La singularidad de estos sonidos es tal que, según Saporiti (1947:434), el nombre vulgar de la especie deriva de una onomatopeya inspirada en su característico grito. Este autor también destaca el comportamiento vigilante y desconfiado del Chajá, así como su predisposición a respuestas defensivas o agresivas.
Ambos sexos participan activamente en la emisión de llamados, frecuentemente mediante duetos que pueden producirse tanto cuando las aves se encuentran posadas como durante el vuelo (Belton, 1984:477; de la Peña, 2025:86). Según de la Peña (2025:86), mientras uno de los integrantes de la pareja emite un potente “AHA”, el otro intercala un agudo “Ogik”, produciéndose ambas vocalizaciones de manera casi simultánea. De forma similar, Sick (1997:242) describe un intercambio vocal durante el vuelo en el que el macho emite un suave “ta-há” y la hembra responde con un fuerte “TÜ-HÜ”.
Existen diferencias sexuales apreciables en las vocalizaciones. Belton (1984:477) indica que ambos sexos poseen voces muy fuertes y de gran alcance, aunque el macho presenta un tono más grave, mientras que la hembra emite llamados más agudos. Brady (2020), citando a Sick (1997:242) y Belton (1984:477), señala igualmente que las vocalizaciones de la hembra son más débiles o más agudas que las del macho. Según Belton (1984:477), el macho produce un llamado breve, gutural y muy grave, descrito como “oh-WOOOW” o, en ocasiones, “be-SERK”, mientras que la hembra emite vocalizaciones más agudas, representadas como “a-ta-HAAA” o “ta-HAAA”, con una terminación de carácter nasal.
Durante la emisión de los llamados, los individuos suelen adoptar posturas corporales características. Sick (1997:242) menciona que, al vocalizar, levantan y sacuden la cabeza. Asimismo, durante el período de cortejo se registra una vocalización particular, descrita por de la Peña (2025:86) como un fuerte “Uahrrrraa… Uahrraaa”, emitido tanto en tierra como en vuelo.
Entre las distintas onomatopeyas registradas para la especie figuran “graía-gragrá” y “chlarü” (Sick, 1997:242), además de las formas descritas por Belton (1984:477) y recopiladas por Brady (2020). En conjunto, estas vocalizaciones desempeñan un papel fundamental en la comunicación entre los miembros de la pareja y constituyen una de las características más reconocibles del comportamiento de la especie.
ALIMENTACIÓNLa alimentación del Chajá (Chauna torquata) es predominantemente herbívora y se basa principalmente en el consumo de vegetación mediante pastoreo, de manera similar a los patos y gansos (Carboneras, 1992:531; Weller, 1999:77, citado por Brady, 2020). Su dieta está compuesta fundamentalmente por hojas, tallos, semillas, plantas acuáticas, vegetación silvestre y algunos cultivos (Brady, 2020; Salvador et al., 2017:16). La excavación del sustrato para la obtención de alimento es muy poco frecuente (Carboneras, 1992:531), y existen registros de individuos cuya alimentación estuvo constituida exclusivamente por vegetación de marisma (Belton, 1984:479).
Los análisis de contenidos estomacales realizados por Aravena (1928:155) evidenciaron un consumo casi exclusivo de material vegetal, especialmente hojas de especies del género Oxalis (O. corniculat var. purpurascens y O. martiana) y flores de “Mata negra/Yuyo moro” (Conyza bonariensis = Erigeron bonariense), sin hallarse restos de origen animal. De forma concordante, Fiora (1939) menciona el consumo de vegetales, mientras que Salvador et al. (2017:16) registraron hojas y tallos de plantas acuáticas, gramíneas y otras especies propias de ambientes asociados a lagunas y bañados.
Aunque la dieta es esencialmente fitófaga, Zotta (1932:78) encontró en contenidos estomacales restos vegetales con semillas, piedritas y numerosos coleópteros, los cuales probablemente fueron ingeridos de manera accidental durante el pastoreo. En conjunto, la evidencia disponible indica que la especie se alimenta principalmente de vegetación característica de ambientes húmedos, siendo excepcional la presencia de componentes no vegetales en su dieta (Aravena, 1928:155; Zotta, 1932:78; Fiora, 1939; Salvador et al., 2017:16; Brady, 2020).
En relación con el comportamiento alimentario, Belton (1984:479) observó que el 24 de octubre de 1983 cuatro individuos, tras nadar hasta la orilla de una pequeña isla en la marisma de Capivari, realizaron repetidos movimientos de estiramiento de las patas antes de dirigirse hacia terrenos más elevados, aportando información complementaria sobre las actividades asociadas al uso de ambientes de alimentación.
REPRODUCCIÓNEl Chajá establece vínculos de pareja duraderos que pueden mantenerse durante varios años e incluso durante toda la vida (Brady, 2020). Durante el cortejo se observan conductas de acicalamiento mutuo, vocalizaciones en dúo y una exhibición característica en la que los individuos retraen la cabeza hacia atrás hasta tocar la espalda (Stonor, 1939).
La reproducción ocurre principalmente durante la primavera austral, con una mayor frecuencia de posturas entre octubre y noviembre, aunque se han registrado nidos y actividad reproductiva en distintos meses del año (Carboneras, 1992:534; Belton, 1984:478).
La especie nidifica de manera solitaria y territorial (Carboneras, 1992:534; de la Peña, 2025:86). Los nidos se ubican en bañados, lagunas, esteros, represas y otros ambientes palustres, generalmente entre juncales, espadañales o pajonales inundados (Salvador et al., 2017:16; de la Peña, 2025:86). Habitualmente son construidos sobre el suelo, aunque ocasionalmente pueden situarse sobre arbustos (de la Peña, 2025:86). Ambos integrantes de la pareja participan en su construcción (Brady, 2020; Carboneras, 1992:532), elaborando una plataforma de juncos, espadañas, ramas y otra vegetación acuática recolectada principalmente en las inmediaciones, aunque en ocasiones el material puede ser transportado desde mayores distancias (Brady, 2020; Carboneras, 1992:532). Gibson (1880:166) describió el nido como una estructura poco profunda de juncos secos, situada sobre el agua o muy próxima a ella.
La puesta consta generalmente de 3 a 5 huevos, aunque puede variar entre 2 y 7 (Carboneras, 1992:532; Brady, 2020). Los huevos son blancos o blanco-crema, ocasionalmente con manchas tenues, y presentan una cáscara granulada (Stonor, 1939; Gibson, 1880:166; Carboneras, 1992:532). En Córdoba se registraron nidadas de 3 a 5 huevos entre septiembre y enero (Salvador et al., 2017:16).
La incubación es compartida por ambos sexos y se prolonga aproximadamente entre 42 y 46 días (Carboneras, 1992:532, 534; Stonor, 1939). Cuando el nido queda temporalmente desatendido, los adultos pueden cubrir los huevos con material vegetal (Carboneras, 1992:532). El cuidado parental posterior también es ejercido por ambos progenitores, quienes atienden a las crías tras la eclosión (Stonor, 1939).
Los pichones nacen cubiertos por abundante plumón gris amarillento o marrón amarillento, más claro en la región ventral (Stonor, 1939; Carboneras, 1992:532, 534; Gibson, 1880:167). Son nidífugos y abandonan el nido poco después de nacer, permaneciendo siempre próximos a sus padres y siguiéndolos de manera constante (Carboneras, 1992:532; Gibson, 1880:167). Durante las primeras semanas utilizan frecuentemente el agua como refugio entre la vegetación debido a su limitada capacidad de desplazamiento terrestre, aunque son buenos nadadores desde edades tempranas (Carboneras, 1992:532; Stonor, 1939).
Los juveniles empluman entre las 8 y 10 semanas de edad y alcanzan la independencia completa entre las 12 y 14 semanas (Stonor, 1939; Weller, 1967; Carboneras, 1992:532). Sin embargo, Gibson (1880:167) observó que los jóvenes pueden permanecer junto a sus padres aun después de adquirir un plumaje similar al de los adultos y antes de desarrollar plenamente la capacidad de vuelo. En concordancia con ello, Belton (1984:478) observó el 24 de enero de 1979 una pareja acompañada por un pichón de 15 a 20 cm de altura cubierto de plumón pardo amarillento, y capturó el 13 de mayo de 1973 un juvenil de tamaño casi adulto que todavía no podía volar.
En Argentina los registros de nidificación abarcan al menos 11 provincias (Buenos Aires, Córdoba, Corrientes, Entre Ríos, Formosa, Jujuy, La Pampa, Mendoza, Salta, San Luis y Santa Fe), siendo Buenos Aires y Santa Fe las jurisdicciones con mayor cantidad de observaciones documentadas, existiendo registros reproductivos para todos los meses del año (de la Peña, 2025:86). Las posturas registradas oscilan generalmente entre 3 y 5 huevos, aunque existen nidadas de hasta 7 huevos (Weller, 1968; De la Peña, 2013:24).
Registros de nidificación y crías de Chauna torquata en Argentina
Provincia Localidad Registro Referencias Buenos Aires Buenos Aires (sin localidad precisa) Nido Stenberg (1869:278) Buenos Aires Provincia de Buenos Aires Posturas en junio y octubre Durnford (1876:162) Buenos Aires Cabo San Antonio, Partido de la Costa Nidos con huevos; posturas en marzo, mayo, junio y agosto Gibson (1880:166); Gibson (1920) Buenos Aires Estancia El Espartillar, General Paz Pollito; nido parasitado por Heteronetta atricapilla Holland (1890:427); Holland (1892:206) Buenos Aires Ajó (Los Yuglases/Los Ingleses) Nido; octubre de 1908 y febrero de 1909 Grant (1911:342) Buenos Aires Zelaya Nido Pereyra (1923:174) Buenos Aires Rosas, Estancia El Toro (Las Flores) Nidificación de septiembre a marzo Daguerre (1922:264) Buenos Aires Cacharí Nido con huevos (14-XI-1919) Smyth (1927:13) Buenos Aires Estancia Cari Lauquen, General Lavalle Nido Gavio (1944:495) Buenos Aires Zoológico de Buenos Aires Posturas en abril, junio, septiembre, octubre y diciembre Saporiti (1947:434) Buenos Aires Zoológico de Buenos Aires 3–6 huevos Haedo Rossi (1970) Buenos Aires Provincia de Buenos Aires Cuatro nidos observados el 01-X-1953 Zuberbühler (1971) Buenos Aires Cabo San Antonio, Partido de la Costa 18 nidos; 4–7 huevos Weller (1968) Buenos Aires Otamendi, Campana Tres nidos; huevos en febrero y pichones en mayo Babarskas et al. (2003:69) Buenos Aires Parque Plaza Montero, Las Flores Nido entre juncos Tajer et al. (2007:7) Buenos Aires General Lavalle Nido parasitado por Heteronetta atricapilla Lyon & Eadie (2013) Buenos Aires Benito Juárez Nido (27-IX-2014) Torres (2014) Buenos Aires Magdalena Cuatro nidos con huevos (1938–1941) Earnshow en Salvador (2016a) Buenos Aires General Madariaga Nido con 4 huevos (13-X-2017) Paulos (2017) Buenos Aires Sin localidad precisa Nido Pagano en Salvador (2016a) Córdoba Morteros, San Justo Nido con 3 huevos (02-I-1982) De la Peña (2013:24) Córdoba Laborde, Unión Tres nidos con 4–5 huevos Nores e Yzurieta Salvador (2016a) Córdoba Departamento San Martín Cinco nidos con 4–5 huevos Salvador y Salvador en De la Peña (2016a) Córdoba Sin localidad precisa Nido Salvador et al. (2017) Corrientes Mburucuyá Dos nidos con 1 y 4 huevos De la Peña (2013:24) Corrientes Esteros del Iberá Nido con 5 huevos (20-VII-2013) Villalba (2013) Corrientes Concepción del Yaguareté Corá Nido con 5 huevos (08-IX-2017) Oscar (2017) Entre Ríos Ceibas, Islas del Ibicuy Nido (18-IX-2011) Lamela (2011) Entre Ríos Gualeguaychú Nido (20-VI-2016) Velázquez (2016) Formosa Reserva/Estancia El Bagual, Laishi Nidos de mayo-julio y octubre-diciembre; 3–5 huevos; crías de mayo-agosto Di Giacomo (2005:230) Jujuy Sin localidad precisa Nido Contino en IML, citado por Salvador (2016a) La Pampa General Pico Nidifica Acevedo et al. (2023) Mendoza Sin localidad precisa Nidifica Lucero (2020a) Salta Paso del Toro, Rosario de Lerma Un huevo (30-VII-1943) Pierotti (IML) Salta Sin localidad precisa Nido Depino et al. en eBird (2015) San Luis San Felipe, Chacabuco Pareja nidificando en juncal Nellar Ramonella (1993) Santa Fe Aguará Grande, San Cristóbal Dos nidos con 3 y 5 huevos De la Peña (2013:24) Santa Fe Marcelino Escalada, San Justo Nido con 5 huevos De la Peña (2013:24) Santa Fe Llambi Campbell, La Capital Tres nidos con 4–5 huevos De la Peña (2013:24) Santa Fe Calchaquí, Vera Siete nidos; hasta 7 huevos; registros de pichones De la Peña (2013:24) Santa Fe Los Amores, General Obligado Cuatro nidos; 1–3 huevos; pareja con 5 pichones De la Peña (2013:24) Santa Fe Paraje 29 Nido en construcción De la Peña (2013:24) Santa Fe Elisa, Las Colonias Nido con 3 pichones (25-XI-2012) Merlino en De la Peña (2016a) Santa Fe Elisa, Las Colonias Ave incubando (15-IX-2017) Merlino (in litt. 2020) Santa Fe Rincón de Ávila, La Capital Nido con ave incubando De la Peña (inéd. 2017) Santa Fe Ruta 62, oeste de Cayastá, Garay Nido con adulto incubando (10-II-2020) De la Peña (inéd. 2020) -
Chajá | Descripción

© Nick Athanas. Chauna torquata. Santa Isobel, Mato Grosso, Brasil. Julio 2014. Algunos derechos reservados Es un ave de gran tamaño, comparable a un pavo, con una longitud de 85–95 cm y un peso promedio de 3.985 g (de la Peña, 2025:86). Se caracteriza por presentar una cabeza grande con copete o cresta de color gris pizarra oscuro. El cuello es gris y está rodeado en su base por un collar negro, precedido por un anillo o banda blanquecina de plumón; algunos autores también mencionan la presencia de una región desnuda alrededor del cuello, no siempre visible (Sick, 1997:242; Castellanos, 1931:387; Lucero, 2020:26).
El iris es pardo oscuro, mientras que la región periocular desnuda y el área implume de la cara son de color rojo con tonalidades rosadas. El pico es negro, corto y robusto, con una longitud aproximada de 4,5 cm, y el lorum se encuentra desnudo (Castellanos, 1931:387; Sick, 1997:242; Lucero, 2020:26).
La coloración general del cuerpo es grisácea o marrón grisáceo oscuro. El dorso es gris oscuro o pizarra, con reflejos acerados, mientras que el pecho presenta tonalidades grises con las plumas bordeadas de blanco. El abdomen es gris claro a blanquecino (Castellanos, 1931:387; Sick, 1997:242; Lucero, 2020:26).
Las alas poseen remeras primarias y secundarias de color pardo oscuro a negro, con cubiertas gris oscuro cuyos bordes son blanquecinos. La superficie superior de las alas es predominantemente negra, pero exhibe una extensa área blanca visible durante el vuelo. Ventralmente, las cubiertas alares y la base de las primarias son blancas, dando a la superficie inferior del ala una apariencia completamente blanca (Sick, 1997:242; Castellanos, 1931:387; Lucero, 2020:26).
Un rasgo distintivo de la especie es la presencia de dos espolones metacarpianos en cada ala, de color rojizo o anaranjado; el espolón principal alcanza una longitud similar a la del pico, mientras que el segundo es considerablemente menor (Castellanos, 1931:387; Sick, 1997:242; Lucero, 2020:26).
La cola es relativamente corta, de aproximadamente 20 cm de longitud, con timoneras de color pardo oscuro u oscuro uniforme (Castellanos, 1931:387; Sick, 1997:242). Las patas son robustas, rojas, rosadas o rojo anaranjadas, con aproximadamente un tercio de las tibias desnudas y uñas negras (Castellanos, 1931:387; Sick, 1997:242; Lucero, 2020:26).
Plumaje juvenil
Los pichones están cubiertos por un plumón suave, fino y denso, de color amarillo con un ligero tinte verdoso. La región ventral es más pálida, mientras que la dorsal presenta tonalidades parduscas. La cabeza es amarillo ocrácea y el pico plomizo. Presentan iris pardos y patas rojo anaranjadas (Lucero, 2020:26).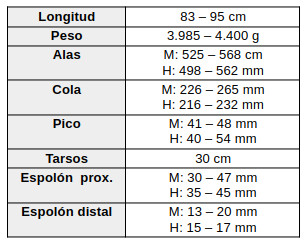
(Rand, 1954:127; Blake, 1977:211; Belton, 1984:477; Carboneras, 1992:534; Brady, 2020; de la Peña, 2025:86) -
Chajá | Taxonomía
Monotípica (Carboneras, 1992:534; Brady, 2020; del Hoyo, 2020:69)
El fósil más antiguo data del Pleistoceno y fue encontrado en Argentina; el fósil tiene aproximadamente 20.000 años de antigüedad (Carboneras 1992:528).
Es muy similar a la Northern Screamer (Chauna chavaria), pero el cuello de la Chauna torquata es casi enteramente negro, y tiene la garganta y los lados de la cara blancos en contraste. Las dos especies no se solapan geográficamente. -
Chajá (Chauna torquata)

El Chajá (Chauna torquata) es el miembro más meridional de una de las familias más características de aves neotropicales, los Anhimidae. Endémica de Sudamérica. Aparece en humedales y pantanos subtropicales a tropicales y puede encontrarse en campos de cultivo.También es una de las aves más grandes que se encuentran en su área de distribución y es fácilmente identificable por su gran tamaño, su cabeza desproporcionadamente pequeña y su plumaje predominantemente gris. Otro rasgo distintivo son los dos espolones óseos de las alas (Brady, 2020)
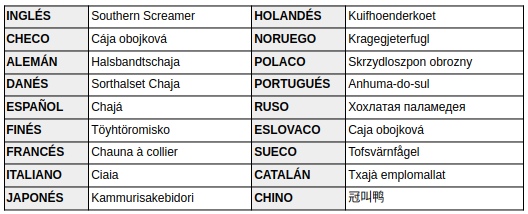
Nombres comunes: Chajá (Argentina); Tachá, Anhupoca, Chaá, Chagá, Chajá, Inhaúma, Saúna, Tachã, tachã-do-sul, tahã (Brasil); Chajá común, Tapacaré (Bolivia); Chaha (Paraguay); También llamado Chahá, Tacat, Tría, Trojoqui, Yajá; En Guaraní: «Chahâ» (Sick, 1997:242; de la Peña, 2025:86; Chébez, 1996:120).
En otros idiomas:
Breves Apuntes Históricos
- F. de Azara (1802). Apuntamientos para la historia natural de los páxaros del Paragüay y Rio de la Plata, Imprenta de la Viuda de Ibarra. Vol III:106 (Chajá)
- Zur Ornithologie Brasiliens: Resultate von Johann Natterers Reisen in den Jahren 1817 bis 1835. Pelzen, A. von, Natterer, J. Wien: A. Pichler’s Witwe & Sohn,1871: 314 (Chauna chavaria)
- Catalogue of the Birds in the British Museum. British Museum (Natural History). Department of Zoology. [Birds]. London, 1874-98. Vol XXVII (Chauna)
- Sclater, PL. Argentine Ornithology. Sclater, Philip Lutley, Hudson, W. H. R.H.Porter, London. (1889); Vol II: 119 (Chauna chavaria?)
-
Familia Anhimidae
La familia Anhimidae comprende aves acuáticas sudamericanas pertenecientes al orden Anseriformes, caracterizadas por una combinación singular de rasgos morfológicos, entre ellos una cabeza relativamente pequeña, cuerpo robusto, espuelas en las alas utilizadas en disputas territoriales y reproductivas, dedos largos escasamente palmeados, sacos dérmicos llenos de aire y la ausencia de procesos uncinados en las costillas. Estas adaptaciones, junto con sus amplias alas aptas para el planeo y sus vocalizaciones roncas y graves, las distinguen de otras aves acuáticas.
Las evidencias moleculares y morfológicas sitúan a los Anhimidae dentro de los Anseriformes (Livezey & Zusi, 2001). Sin embargo, persiste incertidumbre acerca de sus relaciones filogenéticas exactas, ya que podrían constituir el grupo hermano de las demás familias del orden o estar más estrechamente emparentados con Anseranatidae (Donne-Goussé et al., 2002; Eo et al., 2009; Ericson et al., 2006a; Hackett et al., 2008).
Los Anhimidae ocupan diversos humedales tropicales y subtropicales de tierras bajas, incluyendo marismas, sabanas inundables, corredores ribereños y cuerpos de agua extensos. Presentan una dieta predominantemente herbívora basada en hojas, flores, semillas y otras plantas acuáticas, aunque ocasionalmente remueven el barro en busca de alimento.
Su estrategia reproductiva se caracteriza por la monogamia, el cuidado biparental y la aparente formación de vínculos de pareja permanentes. Construyen grandes nidos de vegetación cerca o sobre el agua, donde las hembras depositan entre dos y siete huevos. Ambos progenitores participan en la construcción del nido, la incubación (42–45 días) y la crianza de los polluelos, que alcanzan la independencia entre las 12 y 14 semanas después de la eclosión.
En términos de conservación, solo una de las tres especies reconocidas presenta preocupación inmediata: Chauna chavaria (NT), cuya reducida población, restringida al norte de Colombia y noroeste de Venezuela, muestra una tendencia decreciente.
En Misiones Argentina: (1 Género, 1 especie)
* Género Chauna (2 especies: 1 en Misiones)
– Chauna torquata (Chajá) -
Familia Anatidae