- Acerbo, P. E. (2000). Aves del río Neuquén. Autoridad Interjurisdiccional de Cuencas de los ríos Limay, Neuquén y Negro. 180 pp.
- Acevedo, D. & F. Bruno (2007). Lista de Aves del Parque Don Tomás. Santa Rosa, La Pampa. Editorial L. y M. Gral. Acha. La Pampa. (enlace)
- Acosta, J. C. & F. Murúa (2001). Inventario de la avifauna del Parque Natural Ischigualasto, San Juan, Argentina. Nótulas Faunísticas. Segunda Serie Nº 3: 1-4.
- Acreche, N., Núñez, H.A. & M.V. Albeza (1998).Vulnerabilidad de la avifauna del Parque Nacional Los Cardones, Salta, Argentina. Revista de Biología Tropical 46(3): 811-816. (enlace)
- Administración de Parques Nacionales. S.I.B. Ficha: Coragyps atratus; Recuperado el 10/01/2020 de https://sib.gob.ar/especies/coragyps-atratus
- Administración de Parques Nacionales (2017). Plan de Gestión. Parque Nacional Laguna Blanca. (Revisor del documento: Chehebar, C.).; 222 pp. (enlace)
- Alabarce, E. A., C. Laredo, R. Vides & M. Lucero (1990). Análisis de la estructura de una comunidad de aves en la localidad de Las Juntas, provincia de Catamarca. Estudio preliminar. Acta Zool. Lilloana, 39: 45-52. (enlace)
- Albert, L. A., C. Barcenas, M. Ramos, & E. Iñigo (1989). Organochlorine pesticides and reduction of eggshell thickness in a Black Vulture –Coragyps atratus- population of the Tuxtla Valley, Chiapas, Mexico. In Raptors in the Modern World. pp. 473–475. (enlace)
- Alonso, J. M., & Ronchi-Virgolini, A. L. (2008). Avifauna del Parque Nacional Pre-Delta, Entre Ríos, Argentina. Cotinga, 29(1), 126-134. (enlace)
- Alvarenga, H. M. F. (1985). Notas sobre os Cathartidae (Aves) e descrição de um novo gênero do cenozóico brasileiro. Anais da Academia Brasileira de Ciências 57:349–357. (enlace)
- Antelo, C. M. (1999). Análisis del impacto de las plantaciones de Pinus elliotti sobre la avifauna de Yungas (Tucumán-Argentina). Tesis de Maestría U.N.C., Centro de Zoología Aplicada.
- Antelo, C. M., Z. J. Brandán, C. I. Navarro & N. L. Marigliano (2006). Avifauna de un sector de bosque en Las Tipas (Tucumán-Argentina). Acta Zoológica Lilloana. 50 (1): 41-46. (enlace)
- Antelo, C. M. & Z. J. Brandán Fernández (2013). Las aves no Passeriformes de Tucumán, Argentina. Miscelanea 132. Fundación Miguel Lillo, Argentina. (enlace)
- Antelo, C. M. & C. I. Navarro (2017). Avifauna de un camino rural de montaña en las Yungas de Tucumán (Argentina). Acta Zoológica Lilloana. 61 (1): 3-14. (enlace)
- Anziano, A. (1962). Aves observadas, asi como coleccionadas en el Parque Nacional Nahuel Huapi. (S.I.B.)
- Apochian, C. (2000). Lista adicional de aves observadas durante la realización del Censo Neotropical de Aves Acuáticas. Secc. Río Grande. P.N. Los Alerces. (enlace)
- Aramburú, R. (1983). Lista preliminar de la avifauna de la Laguna Llancanello. Informe inédito, 12 pp
- ArgentAvis (s.f.) Ficha: Jote Cabeza Negra (Coragyps atratus); Recuperado 19/01/2021 de http://argentavis.com.ar/2012/sitio/especie_co.php?id=954
- Athanas, N., Greenfield, P. J., Campbell, I., Cervantes-Daza, P., Spencer, A. & S. Woods (2016). Birds of Western Ecuador. Princeton University Press; 430 pp.
- Aubry, Y., & R. Yank (1993). Quebec region. American Birds 47 (3):393. (enlace)
- Avise, J. C, Nelson, W. S., Sibley, C. G. (1994). DNA sequence support for a close phylogenetic relationship between some storks and New World vultures. Proceedings of the National Academy of Science 91: 5173-5177. (enlace)
- Babarskas, M., J. O. Veiga & F. C. Filiberto (1995). Inventario de Aves del Parque Nacional El Rey, Salta, Argentina. Monografía Especial L. O. L. A. Buenos Aires; Nº 6, 44 pp.
- Babarskas, M. & J. C. Chebez (1999). Notas breves sobre aves de la Argentina y países vecinos. Nuestras Aves 39: 12-14.
- Babarskas, M., E. Haene & J. Pereira (2003). Aves de la Reserva Natural Otamendi. En Haene, E. & J. Pereira (Eds.) Temas de Naturaleza y Conservación. Monografía de aves argentinas 3: 47-113. Aves Argentinas/AOP. Buenos Aires. (enlace)
- Balabusic, A. M., et al. (2001). Plan de Manejo del Parque Nacional Talampaya. Fase II. Administración de Parques Nacionales; 62 pp. (enlace)
- Baliño, J. (1984). Aves del Parque Nacional El Palmar, lista sistemática y otras contribuciones a su conocimiento. Rev. Mus. Arg. Cs. Nat. Zool. 13: 499-511.
- Ballejo, F., & L. J. M. De Santis (2013). Dieta estacional del Jote Cabeza Negra (Coragyps atratus) en un área rural y una urbana en el noroeste patagónico. El Hornero 028 (01):007–014. (enlace)
- Bang, B. G. (1972). The nasal organs of the Black and Turkey vultures: a comparative study of the Cathartid species Coragyps atratus atratus and Cathartes aura septentrionalis. Journal of Morphology 115:153–184. (enlace)
- Bang, B. G., & S. Cobb (1968). The size of the olfactory bulb in 108 species of birds. Auk 85:55–61 (enlace)
- Barnett, J.M., & Pearman, M. (2001). Annotated checklist of the birds of Argentina. Lynx Edicions. 164 pp.
- Barrionuevo, C. A., Julio, L. F. & Carma, M. I. (2002). Determinación de la Composición Especifica y Trófica de la Fauna Ornitológica en el Norte del Valle Central, Dto. Capital, Provincia de Catamarca. Congreso Regional de Ciencia y Tecnología NOA 2002. Secretaría de Ciencia y Tecnología, Universidad Nacional de Catamarca (enlace)
- Bauni, V., R. E. Carini, S. Bogan, J. M. Meluso, T. N. Abad, M. Homberg & A. Giacchino (2017). Inventario de vertebrados del “Paisaje protegido Delta Terra”, primera sección de islas del delta de Tigre, provincia de Buenos Aires, Argentina. Nótulas Faunísticas. Segunda Serie. 210: 1-11. (enlace)
- Beddard, F. E. (1898). The structure and classification of birds. London: Longmans, Green and Co. (enlace)
- Bechard, M. J., Sarasola, J. H. & Helbig, A. J. (2004). Morphometric Measures of male and female Spot‑winged Falconets- Spiziapteryx circumcinctus- sexed using PCR Amplification Methods. Raptors Worldwide. BirdLife Hungary, Budapest. (enlace)
- Belenguer, C. & S. M. Zalba (1997). Primera cita de nidificación del Jote de cabeza negra (Coragyps atratus ) para Buenos Aires. Hornero 014 (04): 247-248. (enlace)
- Beletzky, A. (1979a). Informe de ascenso al Cerro Millaqueo, Parque Nacional Nahuel Huapi. Nota No. 247.
- Beletzky, A. (1979b). Lista estacional de aves, Isla Victoria, P.N. Nahuel Huapi. (S.I.B.)
- Beletzky, A. (1979c). Observación de aves en el P.N. Los Alerces, mes de mayo de 1979. (S.I.B.)
- Beletzky, A. (1981). Lista de aves, Seccional Traful, Parque Nacional Nahuel Huapi. (S.I.B.)
- Beletzky, A. (1984a). Informe de recorridas a la zona del Río Ñirihuau (costa de lago) y Ea. Fortín Chacabuco. P.N. Nahuel Huapi. Nota No. 1570. (S.I.B.)
- Beletzky, A. (1984b). Transecta Costa Sur Lago Traful, lista de flora avistada. Parque Nacional Nahuel Huapi. (S.I.B.)
- Belton, W. (1984). Birds of Rio Grande do Sul, Brazil. Part I. Rheidae through Furnariidae. Bulletin of The American Museum of Natural History; Vol. 178, article 4: 369-636 (enlace)
- Beltzer, A. H. (1981). Lista preliminar de las aves del departamento La Paz (Entre Ríos, Argentina). I. No Passeriformes. Historia Natural 2: 53-56.
- Beltzer, A. H. (1986). Estudio preliminar de la avifauna de la Laguna del Cristal (Cuenca del río Saladillo, Santa Fe, Argentina). Historia Natural 8: 65-74.
- Beltzer, A. H., P. A. Collins & M. A. Quiroga (2006). Atlas ornitogeográfico de la provincia de Entre Ríos. Serie Climax Nº 13. Ceride-Conicet. Santa Fe
- Bendire, C. E. (1892). Life histories of North American birds with special reference to their breeding habits and eggs, with twelve lithographic plates. U.S. National Museum Special Bulletin 1. (enlace)
- Bent, A. C. (1937). Life Histories of North American Birds of Prey. Order Falconiformes. Part I. US National Museum Bulletin 167. (enlace)
- Bertolini, M. P. (1992). Listado de aves del Pasaje Palos Blancos, Ledesma, Jujuy. Inf. Inéd. APN-DTRNO. 2 págs. (S.I.B.)
- Bianchini, M. (2017). Contribuciones al conocimiento de la nidificación de aves en la Patagonia, Argentina. Historia Natural 7:69–83. (enlace)
- Biancucci, L. (1995). La comunidad de aves de la Pampa de Achala. Variación durante el ciclo anual. Cátedra de Diversidad Animal II, FCEF y N, UNC – UNC. 27 pp. (S.I.B.)
- BirdLife International (2020); Ficha: Coragyps atratus. Recuperado de http://www.birdlife.org el 07/10/2021.
- Blake, E. R. (1977). Manual of Neotropical Birds. Vol. 1. Spheniscidae (Penguins) to Laridae (Gulls and Allies). The University of Chicago Press, Chicago & London; 724 pp
- Blendinger, P. G. & M. E. Alvarez (2002). Ensamble de aves de los bañados de Carilauquen (Laguna Llancanello, Mendoza, Argentina) consideraciones para su conservación. Hornero 17: 71-83. (enlace)
- Blendinger, P. G., P. Capllonch & M. E. Alvarez (2004). Abundance and distribution of raptors in the Sierra de San Javier Biological Park, Northwestern Argentina. Ornitología Neotropical. 15: 501-512. (enlace)
- Blendinger, P. G. (2005). Especies de aves observadas en la Reserva Nacional El Nogalar, enero-marzo 2005. Proyecto «Dinámica del reclutamiento de Pino del Cerro (Podocarpus parlatorei): bases ecológicas para su manejo en bosques de montaña del noreste argentino». Informe inéd. a APN-DRNO. 4 pp. (S.I.B.)
- Bodrati, A., Casañas, H. & A. Pietrek (2000). Resultados del primer relevamiento ornitológico del Parque Provincial “Pampa del Indio” (Dto. Lib. Gral. San Martín, Chaco). AOP. Inf. Inéd. Buenos Aires.
- Bodrati, A. & Cockle, K. (2003). Lista de las aves registradas en Villa Unión, Banda Florida y el P.N. Talampaya, Provincia de La Rioja, entre el 25 y 30 de junio de 2003, 3 pp. (enlace)
- Bodrati, A., Mérida, E., Bodrati, G., & Sierra, E. (2006). Avifauna del talar de Vuelta de Obligado y sus ambientes contiguos. San Pedro, Pcia. de Buenos Aires, Argentina. E. Mérida & J. Athor J (Eds.) Talares bonaerenses y su conservación, 117-124. (enlace)
- Bodrati, A., K. Cockle, J. M. Segovia, I. Roesler, J. I. Areta & E. Jordan (2010). La avifauna del Parque Provincial Cruce Caballero, Provincia de Misiones, Argentina. Cotinga 32: 41-64. (enlace)
- Bodrati, A., J. I. Areta & E. White (2012). La avifauna de la Posada y Reserva Puerto Bemberg, Misiones, Argentina. Nuestras Aves 57: 63-80. (enlace)
- Bonkewitzz, A. N. (1990). Fauna de vertebrados de la laguna Guatraché. Universidad Nacional del Sur. Dto. de Biología. Bahía Blanca, 103 pp.
- Braco, M. (1982). Lista de vertebrados observados en Seccional Falkner, Parque Nacional Nahuel Huapi. Nota No. 467. (S.I.B.)
- Brandán, Z. J. (2005). Avifauna de un sector del Bosque de transición Tucumano, en diferentes condiciones de uso. Acta Zoológica Lilloana, 49 (1-2): 3-8. (enlace)
- Brandán, Z. J., N. L. Marigliano, C. I. Navarro & C. M. Antelo (2006). Estructura y composición específica de la comunidad de aves de un área suburbana de la provincia de Tucumán, Argentina. Acta Zoológica Lilloana, 50 (1-2): 47-53 (enlace)
- Brandán, Z. & C. Antelo (2009). Composición y variaciones estacionales de la avifauna de un ambiente antropizados (San Pablo, Tucumán, Argentina). Acta Zoológica Lilloana, 53 (1-2): 98-103. (enlace)
- Brandán, Z. J. & C. I. Navarro (2009). Lista actualizada de las aves de la provincia de Tucumán. Fundación Lillo. Tucumán. (enlace)
- Brandán Fernández, Z. J. & N. L. Marigliano (2019). Lista comentada de las aves del río Nío/Tajamar, Dto. Burruyacu, Tucumán, Argentuna. Nótulas Faunísticas. Segunda Serie. 275: 1-3. (enlace)
- Brandolín, P., R. Martori & M. Ávalos (2007). Variaciones temporales de los ensambles de aves de la Reserva Natural de Fauna Laguna La Felipa (Córdoba, Argentina). Hornero 022 (01): 001-008. (enlace)
- Brandolin, P., Blendinger, P., Ramírez, R., Márquez, J., Miguel, Á. & Cantero, J. (2016). Birds of a wildlife reserve in the South American Pampa (Córdoba, Argentina). Check List, 12, 1. (enlace)
- Brooks, T. M., Barnes, R., Bartrina, L., Butchart, S. H. M., Clay, R. P., Esquivel, E. Z., Etcheverry, Ni. I., Lowen, J. C. & J. Vincent (1993). Project CANOPY’92: Final Report. (enlace)
- Brown, L. H., & D. Amadon (1968). Eagles, Hawks, and Falcons of the World. McGraw-Hill, New York, NY, USA; 945 pp.
- Brown, W. H. (1976). Winter population trends in the Black and Turkey Vultures. American Birds 30:909–912. (enlace)
- Bruno, F., Güller, R., Güller, P. & Erro, M. (2008). Aves observadas y consideraciones sobre el estado actual del AICA Victorica, Provincia de La Pampa, Argentina. Informe interno, Aves Argentinas /AOP, 6 pp.
- Brunton, D. F., & T. Pratt (1986). Sightings of a Black-throated Sparrow, Amphispiza bilineata, and a Black Vulture, Coragyps atratus, in British Columbia. Canadian Field-Naturalist 100:256–257. (enlace)
- Buckley, N. J. (1996). Food finding and the influence of information, local enhancement, and communal roosting on foraging success of North American vultures. Auk 113:473–488. (enlace)
- Buckley, N. J. (1997). Experimental tests of the information-center hypothesis with Black Vultures (Coragyps atratus) and Turkey Vultures (Cathartes aura). Behavioral Ecology and Sociobiology 41:267–279. (enlace)
- Buckley, N. J. (1998). Interspecific competition between vultures for preferred roost positions. Wilson Bulletin 110:122–125. (enlace)
- Buckley, N. J. (2020). Black Vulture (Coragyps atratus), version 1.0. In Birds of the World (A. F. Poole and F. B. Gill, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.blkvul.01
- Budin, E. (1931). Lista y notas sobre aves del N. O. Argentino (Prov. de Jujuy). Hornero 4: 401-411. (enlace)
- Bullock, D. S. (1956). Vultures as disseminators of anthrax. Auk 73:283–284. (enlace)
- Burgos, F. G., J. L. Baldo & F. M. Cornell (2009). Lista de las aves de la Provincia de Jujuy, Argentina. Secretaría de Turismo y Cultura de Jujuy. S. S. de Jujuy. (enlace)
- Cabido, M., Anton, A., Cabrera, M., Cingolani, A., Diaz, S., Di Tada I. & Falczuk V. (2003). Informe Final de Linea de Base de la Biodiversidad y Programa de Monitoreo del P.N. Quebrada del Condorito y Reserva Hidrica Provincial Pampa de Achala. (S.I.B.)
- Camperi, A. R. (1998). Avifauna Andinopatagónica, lista comentada de especies. Physis Sec. C. 56 (130-131): 33- 46
- Camperi, A. R. & C. A. Darrieu (2000). Avifauna de Catamarca: Lista comentada de especies (No passeriformes). Physis Sec. C. 58 (134-135): 67-78.
- Camperi, A., Darrieu, C., Grilli, P., & Burgos, F. (2012). Avifauna de la provincia de Jujuy, Argentina: lista de especies (no Passeriformes). Acta zoológica lilloana, 56 (1-2): 82-140. (enlace)
- Canavelli, S., M. E. Zaccagnini, J. Torresín, N. Calamari , M. de la Paz Duccomun & P. Capllonch (2004). Monitoreo extensivo de aves en el centro-sur de Entre Ríos. Insugeo, Miscelánea 12: 349-362. (enlace)
- Capllonch, P. (1998). La avifauna de los bosques de Transición del Noroeste Argentino. Tesis doctoral, Facultad de Ciencias Naturales e Instituto M. Lillo, Universidad Nacional de Tucumán, 157 pp.
- Capllonch, P., Lobo, R., Ortiz, D. & R. Ovejero (2005). La avifauna de la selva de galería en el noreste de Corrientes, Argentina: Biodiversidad, patrones distribución y migración. Insugeo, Miscelánea, 14: 483-498. (enlace)
- Capllonch, P. (2007). Aves de Tafí del Valle. Tucumán. Argentina. (enlace)
- Capllonch, P., Ortiz, D., & Soria, K. (2008). Importancia del litoral fluvial argentino como corredor migratorio de aves. Insugeo, Miscelánea: 17: 107-120. (enlace)
- Capllonch, P., D. Ortiz & I. Ferro (2011). Notas sobre la avifauna de las Cumbres Calchaquíes, Tucumán, Argentina. Acta Zoológica Lilloana 55 (1): 50-63. (enlace)
- Capmourteres, V., V. Bauni, J. M. Meluso, S. Bogan & M. Homberg (2015). Aves de la Reserva Natural Campo San Juan, Misiones: descripción e implicancias para su conservación. Nótulas Faunísticas. Segunda Serie. Nº 171: 1-14. . (enlace)
- Carma, M. I. (2007). Relevamiento de aves de la provincia de Catamarca. (1º Ed.). Consejo Federal de Inversiones, CABA, Argentina, 2009; 500 pp. (enlace)
- Casares, J. (1944). Aves de Estanzuela. San Luis. Hornero 008 (03): 379-463. (enlace)
- Castellanos, A. (1932). Aves del valle de los Reartes (Córdoba). Hornero 005 (01): 1-40. (enlace)
- Cebolla-Badie, M. (2000). El conocimiento mbya-guaraní de las aves. Nomenclatura y Clasificación. Suplemento Antropológico. Universidad Católica. Revista del Centro de Estudios Antropológicos. Vol XXXV, Nº2. ISSN 0378-9896. (enlace)
- Ciarmiello, D., Fabri., S. & Gil, G. (2005). Campaña de prospección del estado de conservación de un sector del río Iguazú Superior (S.I.B.)
- Cingolani, A. & V. Falczuk (2003). Elaboración Plan de Manejo Integrado del P.N. Quebrada del Condorito y la Reserva Hídrica Provincial de Achala. Cuarto informe de Consultoría; 222pp.
- Clapp, R. B., M. K. Klimkiewicz, & J. H. Kennard (1982). Longevity records of North American birds: Gaviidae through Alcidae. Journal of Field Ornithology 53:81–208. (enlace) (enlace)
- Claramunt, S. & Cuello, J. P. (2004). Diversidad de la biota uruguaya. Aves. An. Mus. Hist. Nat. y Antropología, 10(6), 1-76. (enlace)
- Clark, R. G., & R. D. Ohmart (1985). Spread-winged posture of Turkey Vultures: single or multiple functions? Condor 87:350–355. (enlace)
- Clay, T. (1951). The mallophaga and relationships within the Falconiformes. Ibis 93:628. (enlace)
- Coconier, E. G., López Lanús, B., Roesler, I., Moschione, F., Pearman, M., Blendinger, P., Bodrati, A., Monteleone, D., Casañas, H., Pugnali, G. & M. E. Alvarez (2007). Lista comentada de las aves silvestres de la unidad de gestión Acambuco. Pp: 32-103. En Coconier, E. G. (Ed.). Las aves silvestres de Acambuco, Provincia de Salta, Argentina. Temas de Nat. y Cons. Nº 6. Nuestras Aves A.O.P. (enlace)
- Coleman, J. S. J.D. Fraser & C. A. Pringle (1985) Salt eating by Black and Turkey Vultures. Condor 87:291-292. (enlace)
- Coleman, J. S. & J. D. Fraser (1986). Predation on Black and Turkey vultures. Wilson Bulletin 98:600–601. (enlace)
- Coleman, J. S. & Fraser, J. D. (1987). Food habits of Black and Turkey vultures in Pennsylvania and Maryland. Journal of Wildlife Management; 51: 733–739. (enlace)
- Coleman, J. S., J. D. Fraser, & P. F. Scanlon (1988). Hematocrit and protein concentration of Black Vulture and Turkey Vulture blood. Condor 90:937–938. (enlace)
- Coleman, J. S., & J. D. Fraser (1989a). Age estimation and growth of Black and Turkey Vultures. Journal of Field Ornithology 60:197–208. (enlace)
- Coleman, J. S., & J. D. Fraser (1989b). Habitat use and home ranges of Black and Turkey vultures. Journal of Wildlife Management 53:782–792. (enlace)
- Coleman, J. S., & J. D. Fraser (1990). Black and Turkey vultures. In Proceedings of the Southeast Raptor Management Symposium and Workshop. National Wildlife Federation, Washington, DC, USA. pp. 78–88.
- Comisso, E. (2012). Informe de especies de fauna observadas en el Parque Nacional Los Alerces 2007-2012. (S.I.B.)
- Contaldi, M. D. M. B., Bildstein, K. & Fernandez, G. J. (2019). Ecología del jote cabeza negra (Coragyps atratus) en Patagonia. (enlace)
- Contino, F. (1980). Aves del Noroeste Argentino. U.N.S., Secretaría de Estado de Asuntos Agrarios, Dirección General de Recursos Naturales Renovables, 135 pp.
- Contreras, J. R. (1977). La avifauna del Valle del río Collón Curá, provincia del Neuquén. Idia. Suplemento 34: 250-270.
- Contreras, J. R. (1979). Lista faunística preliminar de los vertebrados de la Reserva Ecológica de Ñacuñan, Cuaderno Técnico (IADIZA) 1: 39-47.
- Contreras, J., Roig, V. & Giai, A. (1980). La avifauna de la Cuenca del Río Manso Superior y la orilla sur del Lago Mascardi, P.N. Nahuel Huapi, Provincia de Río Negro Historia Natural. Vol.1. (8):41-48. 1980
- Contreras, J. R. (1981). Lista preliminar de la avifauna correntina. I. No Passeriformes. Historia Natural 2: 21-28.
- Contreras, J. R. (1987). Lista preliminar de la avifauna de la provincia de Formosa, República Argentina. Historia Natural VII (4): 33-52.
- Contreras, J. R., L. M. Berry, A. O. Contreras, C. C. Bertonatti & E. E. Utges (1990). Atlas ornitogeográfico de la provincia del Chaco. República Argentina. I. No passeriformes. Cuad. Téc. Félix de Azara Nº 1. 164 pág.
- Contreras, J. R., F. Agnolin, Y. E. Davies, I. Godoy, A. Giacchino & E. Ríos (2014). Atlas ornitogeográfico de la provincia de Formosa, República Argentina. 1º Ed. Ciudad Autónoma de Buenos Aires: Fundación de Historia Natural Félix de Azara. 450 pp. (enlace)
- Coria, O. R., J. Heredia, R. Torres, J. Lima, P. Araujo, H. Zerda, M. Palacio & E. Roger (2017). Lista de aves de la sierra de Guasayán, Santiago del Estero, Argentina. Nótulas Faunísticas. Segunda Serie. 212: 1-14. (enlace)
- Chalukian, S. C. (1989). Relevamiento ecológico del Parque Nacional El Rey: Fauna. Informe de beca interna. Administración de Parques Nacionales. 100 pp. (S.I.B.)
- Chalukian, S., de Bustos, S., Saravia, M., Lizárraga, R. L., Serapio, R., Sola, M. F. & M. Romano (2002). Recategorización de Áreas Protegidas en el Corredor Transversal Sur, Salta, Argentina. Informe final. Secretaría de Medio Ambiente y Desarrollo Sustentable de la Provincia de Salta (SeMADeS) y Wildlife Conservation Society (WCS). 63 pp. Localización física del documento: DRNO (S.I.B.)
- Chatellenaz, M. L. (2004). La avifauna del bosque de Quebracho colorado y Urunday del noroeste de Corrientes, Argentina. Facena, 20: 3-12. (enlace)
- Chatellenaz, M. L. (2005). Aves del Valle del Río Paraná en la Provincia del Chaco, Argentina: riqueza, historia natural y conservación. Insugeo, Miscelánea 14: 527-550. (enlace)
- Chatellenaz, M. L., P. Cano, C. Saibene & H. A. Ball (2010). Inventario de las aves del Parque Nacional Mburucuyá (provincia de Corrientes, Argentina). Acta Zool. Lilloana 54: 139-160. (enlace)
- Chebez, J. C., N. R. Rey, M. Babarskas & A. G. Di Giacomo (1998). Las aves de los Parques Nacionales de la Argentina. Monografía Especial L.O.L.A. Nº 12; 126 pp. Bs.As.
- Chebez, J. C. (1996). Fauna misionera, catálogo sistemático y zoogeográfico de los vertebrados de la provincia de Misiones (Argentina). Monografía Nº 5. Págs. 320. L. O. L. A. Bs.Aires.
- Chebez, J., Bosso, A., Heinonen, S., Liva, A. & Chavez, H. (2001). Aves del Parque Natural Municipal Dr. Luis Honorio Rolón. Puerto Iguazú, Misiones.
- Chebez, J. C. (2005). Guía de las Reservas Naturales de la Argentina. Noroeste. Ed. Albatros, Buenos Aires; 288 pp.
- Chebez, J. C. & N. S. Morandeira (2007). Parque Nacional El Palmar. En A. S. Di Giacomo (editor). Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 168-170. Temas de Naturaleza y Conservación 5. Aves Argentinas/Asociación Ornitológica del Plata. Buenos Aires. (enlace)
- Chehebar, C., E. J. Ramilo & S. A. Mazzuchelli (1992). Parque Nacional Laguna Blanca. Plan General de Manejo. Documento preliminar para discusión. Delegación técnica regional. Informe inédito. Administración Parques Nacionales. (enlace)
- Chehebar, C. & E. J. Ramilo (1992). Fauna del Parque Nacional Nahuel Huapi. Administración de Parques Nacionales y Asociación Amigos del Museo de la Patagonia “Francisco Pascasio Moreno”, San Carlos de Bariloche, 38 pp.
- Chesser, R. T., S. M. Billerman, K. J. Burns, C. Cicero, J. L. Dunn, A. W. Kratter, I. J. Lovette, N. A. Mason, P. C. Rasmussen, J. V. Remsen, Jr., D. F. Stotz, & K. Winker (2020). Check-list of North American Birds (online). A.O.S.(enlace)
- Chester, S. (2008). A Wildlife Guide to Chile. (1º Ed.). Published by Princeton University Press, 41 William Street, Princeton, New Jersey 08540. 392 pp.
- Christie, M. (1984). Informe de evaluación del interés biológico del area de Villa Traful. Actual Comisión de fomento, antes: Parque Nacional Nahuel Huapi. (S.I.B.)
- Christie, M. I., E. J. Ramilo & M. D. Bettinelli (2004) Aves del noroeste patagónico. Atlas y guía. L. O. L. A. pp. 328
- Cid, D. F. & Caviedes-Vidal, E. (2005). Avifauna da Represa “La Florida” (San Luis, Argentina). Atualidades Ornitológicas N.125:10. (enlace)
- Daciuk, J. (1968). Estación biológica de Puerto Radal, Isla Victoria, para la investigación de la vida silvestre. P.N. Nahuel Huapi. Exp. No. 5814/68. (S.I.B.)
- Dardanelli, S., Serra, D. A., & Nores, M. (2006). Composición y abundancia de la avifauna de fragmentos de bosque de Córdoba, Argentina. Acta zoológica lilloana, 50(1-2), 71-83. (enlace)
- Darrieu, C.A. & A.R. Camperi (2001) Nueva lista de las aves de la provincia de Buenos Aires. Cobiobo Nº 3-Probiota No 2. Sec. de Política Ambiental. UNLP. Buenos Aires. (enlace)
- Darrieu, C. A., A. R. Camperi, & S. Imberti (2008). Avifauna (no Passeriformes) of Santa Cruz province, Patagonia (Argentina): annotated list of species. Revista del Museo Argentino de Ciencias Naturales 10:111–145. (enlace)
- Darrieu, C. A., A. R. Camperi, J. J. Maceda & F. Bruno (2011). Avifauna de la provincia de La Pampa, Argentina: lista de especies (no passeriformes) Acta Zoologica Lilloana. 55: 64 – 108. (enlace)
- Decker, M. D., P. G. Parker, D. J. Minchella, & K. N. Rabenold (1993). Monogamy in Black Vultures: genetic evidence from DNA fingerprinting. Behavioral Ecology 4:29–35. https://doi.org/10.1093/beheco/4.1.29
- De la Peña, M. R. (1987). Nidos y huevos de aves argentinas. (Ed. del autor). Santa Fe, Argentina.
- De la Peña, M. R. (1999). Aves argentinas. Lista y distribución. L.O.L.A. (Literature of Latina America). Buenos Aires. 244 pp.
- De la Peña, M. R. (2001). Observaciones de campo en la alimentación de las aves. Rev. FAVE. Vol. 15 N° 1: 99- 107. (enlace)
- De la Peña, M. R. (2006) Lista y distribuciónde las aves de Santa Fe y Entre Ríos. Monografía Nº 15; L.O.L.A. (Literature of Latin America), Buenos Aires; 138 pp.
- De la Peña, M. R., F. C., Laene Silva, R., Capuccio, G. & Bonín, L. M. (2009) Aves del Río Uruguay. Guía Ilustrada de Especies del Bajo Uruguay y el Embalse de Salto Grande. Comisión Administradora del Río Uruguay. CARU. 247 pp. (enlace)
- De la Peña, M. R. & S. A. Salvador (2010). Manual de la alimentación de las aves argentinas. UNL. (Digital) (enlace)
- De la Peña, M. R. & R. F. Tittarelli (2011a). Guía de aves de La Pampa. Santa Rosa. Gobierno de La Pampa
- De la Peña, M. R. (2011b). Atlas ornitogeográfico de la provincia de Santa Fe. Serie Naturaleza y Conservación No 4. Pág. 1-500
- De la Peña, M. R. (2012). Distribución y citas de aves de Entre Ríos. Serie Naturaleza y Conservación Nº 6. Pág. 1- 206.
- De la Peña, M. R. (2013). Nidos y reproducción de las aves argentinas. Ediciones Biológica. Serie Naturaleza, Conservación y Sociedad N° 8. Santa Fe, Argentina. 590 pp. (enlace)
- De la Peña, M. R. (2015) Aves argentinas (1º Ed.) Santa Fe; Ediciones U.N.L. C.A.B.A.: EUDEBA; Vol 1: 496 pp.
- De la Peña, M. R. (2020). Aves Argentinas. Descripción, Comportamiento, Reproducción y Distribución (Actualización). Comunicaciones del Mus. Prov. de Ciencias. Nat. “Florentino Ameghino” (Nueva Serie) Año 2020 | N° 4 | Pág. 1-255
- del Hoyo, J. (2020). All the Birds of the World. Lynx Edicions, Barcelona.
- De Lucca, E. R. (2016). Reproducción del Jote cabeza negra (Coragyps atratus) en un nido construido por Loros barranqueros (Cyanoliseus patagonus) en un acantilado del litoral patagónico, Argentina. Nótulas Faunísticas 194: 1-6. (enlace)
- De Lucca, E. R. & J. P. De Lucca (2017). Aves de presa diurnas que nidifican en estructuras construidas por el hombre en la Argentina. Nuevos aportes y una revisión. Nótulas Faunísticas. Segunda Serie. 220: 1-10. (enlace)
- de Viana, M. L., Garrido, J. L., Regidor, H., Núñez, A. & R. Acosta (1998). El ambiente biótico de la obra. Pp. 109-142. En: Estudio de Impacto Ambiental. Ruta Provincial No. 33. Tramo El Carril – Payogasta. Sector Piedra de Molino a Cajoncillo y Cajoncillo acceso a Tonco. Dirección de Vialidad, Gobierno de la Provincia de Salta. IRNED – UNSa. 186 pp. (S.I.B.)
- Dieguez, A. & Sijovich, C. (1997). Conteo de aves rapaces (Accipitridae-Falconidae-Cathartidae-Strigidae) en la Reserva Natural Otamendi, Pdo. de Campana, Pcia de Bs. As. Aprona. Boletín Científico, nov. 1997. (enlace)
- Di Giacomo, A. G. (2005). Aves de la Reserva El Bagual. En Di Giacomo, A. G. y S. F. Krapovickas (Eds.). Historia natural y paisaje de la Reserva El Bagual, provincia de Formosa, Argentina. Inventario de la fauna de vertebrados y de la flora vascular de un área del Chaco Húmedo. Temas de Naturaleza y Conservación 4: 201-465. Aves Argentinas/AOP. Buenos Aires. (enlace)
- Echevarria, A. L., Marigliano, N. L., & Chani, J. M. (1998). Composición y variaciones anuales de la diversidad de aves de una localidad de Bosque Chaqueño Serrano (Ticucho, El Cadillal, Tucumán, Argentina). Acta Zoológica Lilloana, 44(1), 207-217. (enlace)
- Echevarría, A. L. & J. M. Chani (1999). Lista de aves del embalse El Cadillal, provincia de Tucumán. Acta Zoológica Lilloana 45 (1): 141-145. (enlace)
- Echevarría, A. L. & J. M. Chani (2000). Estructura de las comunidades de aves acuáticas del embalse El Cadillal, Tucumán, Argentina. Acta Zoológica Lilloana 45 (2): 219-232. (enlace)
- Echevarria, A. L. (2001). Estudios ecológicos de las aves acuáticas del embalse El Cadillal, provincia de Tucumán. Tesis doctoral, Facultad Ciencias Naturales e Instituto M. Lillo, Universidad Nacional de Tucumán, 206 pp.
- Echevarría, A. L. & J. M. Chani (2006). Aves migratorias, la importancia del embalse El Cadillal, (Tucumán, Argentina) como sitio de tránsito e invernada. Acta Zoológica Lilloana 50 (1-2): 97-108. (enlace)
- Echevarria, A., Marano, C. & Fanjul, E. (2008a). Comunidades de aves del embalse Escaba y sus alrededores, Tucumán. Resumen de XII Reunión Argentina de Ornitología, 5 al 8 de marzo 2008, San Martín de los Andes, Neuquén, Argentina. (enlace)
- Echevarría, A. L., C. F. Marano, J. M. Chani & M. C. Cocimano (2008b). Composición de la comunidad de aves del embalse La Angostura, Tafí del Valle, Tucumán. Acta Zoológica Lilloana 52 (1-2): 98-105. (enlace)
- Echevarría, A. L., C. F. Marano, M. C. Cocimano, M. E. Fanjul & M. A. Cormenzana (2014). Composición y variación de la comunidad de aves del embalse El Tunal, Salta, Argentina. Acta Zoológica Lilloana 58 (1): 80-93. (enlace)
- Echevarria, A. L., & Fanjul, M. E. (2016). Estructura, composición y variación estacional de la avifauna del Embalse Escaba (selva montana subtropical), Tucumán, Argentina. Acta zoológica lilloana, 101-115. (enlace)
- Echevarría, A. L., M. E. Fanjul, M. V. Martínez & A. Benavidez (2019). Avifauna de los bañados de Añatuya, área de importancia para la conservación de las aves (AICA) de Santiago del Estero, Argentina. Hornero 34 (2): 43-49. (enlace)
- EcoRegistros (2020). Ficha: Jote Cabeza Negra (Coragyps atratus) (Bechstein, 1783); Recuperado de https://www.ecoregistros.org/ficha/Coragyps-atratus, el 19/01/2020
- Eisenmann, E. (1963). Is the Black Vulture migratory? Wilson Bulletin 75:244–249. (enlace)
- Elias, D. J., & G. D. Valencia (1982). Unusual feeding behavior by a population of Black Vultures. Wilson Bulletin 94:214. (enlace)
- Emslie, S. D. (1988). An early condor-like vulture from North America. Auk 105:529–535. (enlace)
- Ericson, P.G.P., Anderson, C.L.A., Britton, T., Elzanowski, A., Johansson, U.S., Källersjö, M., Ohlson, J. I., Parsons, T. J., Zuccon, D. & Mayr, G. (2006). Diversification of Neoaves: integration of molecular sequence data and fossils. Biology letters, 2(4), 543-547 (enlace)
- Fandiño, B., & Giraudo, A. R. (2010). Revisión del inventario de aves de la provincia de Santa Fe, Argentina. U.N.L. (enlace)
- Fandiño, B. & A. A. Pautasso (2011). Avifauna del AVP La Salamandra, pp.: 62-105. En: Pautasso, A. A. 2011. La fauna y su conservación en los Bajos Submeridionales. Ediciones Biológica. Serie Naturaleza, Conservación y Sociedad No 5. Santa Fe, Argentina, 278 pp. (enlace)
- Fandiño, B., L. A. Leiva, A. A. Pautasso, H. Luna & M. Manassero (2015). Avifauna de las reservas natural manejada “El Fisco” y privada de uso múltiple “Isleta linda”, Santa Fe, Argentina. Nuestras Aves 60: 39-45. (enlace)
- Fandiño, B., J. M. Fernández, M. L. Thomann, R. Cajade & A. B. Hernando (2017). Comunidades de aves de bosques y pastizales en los afloramientos rocosos aislados del Paraje Tres Cerros, Corrientes, Argentina. Revista de Biología Tropical Vol. 65 (2): 535-550. (enlace)
- Fandiño, B. (2020). Inventarios de aves de la Reserva Natural Estricta La Loca y la Reserva Natural Manejada Potrero 7B, en la “Cuña Boscosa”(Chaco Húmedo) de Santa Fe, Argentina. Acta Zoológica Lilloana, 130-147. (enlace)
- Farinaccio, F. M. (2008). Determinación de la estructura del ensamble de aves de un sector de humedal periurbano de la ciudad de Plottier, provincia de Neuquén. Tesis de Grado. Universidad Nacional de Comahue (enlace)
- Fariñas Torres, T., Rios, A., Schiappacasse, E., Mañez, M., Beruhard, J. & Morales, R. (2019). Relevamiento de fauna para la puesta en valor de la Reserva Natural de Usos Múltiples Cerro del Cóndor, departamento Pocho, Córdoba, Argentina. (enlace)
- Fava, G., Acosta, J. C., Laspiur, J. A. & Moreno, M. V. (2012). Avifauna del Valle de Calingasta y sus alrededores, provincia de San Juan, Argentina. Acta zoológica lilloana, 66-81. (enlace)
- Fava, G. A., J. C. Acosta G. & M. Blanco (2017). Efecto de la estacionalidad y lluvias en la avifauna del Chaco Serrano Austral, Argentina. Rev. Biol. Trop. Vol. 65 (3): 953-961. (enlace)
- Feduccia, A. (1996). The Origin and Evolution of Birds. Yale University Press, New Haven, CT, USA.
- Ferguson-Lees, J. & D. A. Christie (2001). Raptors of the World. Christopher Helm, London, United Kingdom.
- Fernandez, J. M., Thomann, L., Fandiño, B., Cajade, R. & Hernando, A. (2021). An annotated checklist of birds of Paraje Tres Cerros, Corrientes province, Argentina. Check List, 17, 415. (enlace)
- Fernández, V., & Borghi, C. E. (2010). Nuevos registros y comentarios sobre la avifauna del noreste de la provincia de San Juan y el Parque Provincial Ischigualasto. (enlace)
- Ferrer, D., R. Olivera, A. Elías, M. Pérez & P. Isola (2019). Lista actualizada de aves registradas en la provincia de Mendoza, Argentina. Historia Natural , Tercera Serie. Vol. 9 (2): 81-106. (enlace)
- Ferreyra, M. & Vidoz, F. (2007). Exploración de la flora, vegetación y fauna altoandinas en el Parque Nacional Lago Puelo. Informe Final. Año 2007. (enlace)
- Fiora, A. (1939). Distribución geográfica de las especies de aves capturadas en la provincia de Jujuy y zonas limítrofes. Physis 16: 272-289.
- Fiori, S. M. & S. M. Zalba (1999). Plan de Manejo de la Reserva Provincial Auca Mahuida, Neuquén. GEKKO Grupo de estudios en Conservación y Manejo. Departamento de biología, Bioquímica y Farmacia, Universidad Nacional del Sur-Bahía Blanca; 260 pp. (enlace)
- Fisher, H. I. (1939). Pterylosis of the Black Vulture. Auk 56:407–410. (enlace)
- Floria, M. P. (2014). Monitoreo de aves en el Parque Nacional Los Alerces. Zona Sur. Proyecto Eólico Futaleufú. Informe semestral ( enero-junio 2014) (S.I.B.)
- Fraga, R. M. (2001). The avifauna of Estancia San Juan Poriahú, Iberá marshes, Argentina: check-list and some natural history notes. Cotinga 16: 81-86. (enlace)
- Frenzel, J. (1891). Uebersicht uber die in der Provinz Córdoba (Argentinien) vorkommenden Vogel. J. F. Orn., 194: 113-127. (enlace)
- Friedmann, H. (1950). The Birds of North and Middle America. Part XI. Cathartidae to Falconidae. U.S. National Museum Bulletin no. 50. Smithsonian Institution, Washington, DC, USA. (enlace)
- Gallardo, J. M. (1987). Observaciones sobre Coragyps atratus (Aves, Cathartidae) en semilibertad. Revista del M. A.C.N., Bernardino Rivadavia. Zoología, 14 (1).
- Garrahan, P. (1980). Aves observadas en la Isla Victoria, PN Nahuel Huapi. (S.I.B.)
- Garrido, O. H. & A. Kirkconnell (2000). Field Guide to the Birds of Cuba. Cornell University Press, Ithaca, NY, USA.
- Gelain, M. A. (1984). La avifauna del Cerro Otto. Nuestras aves 3: 4-5.
- Gelain, M. A. (2001). Lista sistemática de las aves del Municipio de San Carlos de Bariloche, Río Negro, Argentina. Ed. Grupo Naturalista Sur Patagonia.
- Gelain, M. A., L. Sympson & F. F. Vidoz (2003). Aves de Bariloche. Lista comentada de aves del departamento Bariloche, Provincia de Río Negro, Argentina. Libros del Mediodía. San Carlos de Bariloche.
- Gelain, M. A. (2010). Aves de la patagonia argentina: atlas ornitológico de Río Negro. San Juan
- Gelain, M. A. & J. Tolosa (2011a). Aves de la provincia de Río Negro. Lista sistemática. Xolmis CRO 6: 1-37
- Gelain, M. A. & R. Pereyra Lobos (2011b). Lista de aves de la provincia de San Juan, Argentina. Xolmis CRO 3: 1- 12.
- Gelain, M. A. (2014). Lista preliminar de aves del departamento Angaco, provincia de San Juan, República Argentina. Xolmis 35: 1-27.
- Giacchino, H., Posadas, C. & Williams, D. (1980). Informe de la recorrida realizada a la cabecera O. del lago Martin, Seccional Inacayal. Nota No. 263. (S.I.B.)
- Giai, A. G. (1950). Notas de viajes. Por el norte occidental de Santa Fe y por el norte de Misiones. Hornero 9: 121- 164. (enlace)
- Gil, G. (1996). Primer relevamiento expeditivo del Parque Nacional Campo de los Alisos. Inf. Inéd. APN-DTRNO. 36 p gs. (S.I.B.)
- Giraudo, A. R., J. L. Baldo & R. R. Abramson (1993). Aves observadas en el sudeste, centro y este de Misiones (República Argentina), con la mención de especies nuevas o poco conocidas para la provincia. Nótulas Faunísticas 49: 1-13. (enlace)
- Giraudo, L., M. Kufner, R. Torres, D. Tamburini, V. Briguera & G. Gavier (2006). Avifauna del Bosque Chaqueño Oriental en la provincial de Córdoba, Argentina. Ecología Aplicada 5: 127-136. (enlace)
- Girini, J. M. (2014). Informe de avance de proyecto: «Cambios en las poblaciones de aves ante la presencia de visón americano Neovison vison en lagos y lagunas del Parque Nacional Lanín, Neuquén». (S.I.B.)
- Gómez, V. E. (2006). Aves del Parque General San Martín (Mendoza). Distribución y características. Multequina, 15 (2): 81-95, versión on line. (enlace)
- Gómez, A. & F. Vocos (1992). Especies detectadas en el Parque Nacional El Rey (Salta, Rep. Arg.). Inf Inéd. APN-DTRNO. 4 pp. (S.I.B.)
- Gorgoglione, C. E. (2005). Guía de campo para la identificación de las aves del Neuquén. Edición del autor. 3º Edición. Junín de los Andes. 279 p.
- Gorleri, F., M. C. Gorleri & M. Murdoch (2011). Aves de la Reserva de Biosfera Laguna Oca, Formosa, Argentina. Universidad Nacional de Formosa/SPU Programa de Voluntariado Universitario. Ideas Gráficas. Formosa. 158 pp.
- Gorleri F. & Di Giácomo A.G. (2011) Lista sistemática de las aves de la Reserva de Biosfera Laguna de Oca del Río Paraguay, Formosa. Aves argentinas (enlace)
- Greeney, H. F., R. A. Gelis, & R. White (2004). Notes on breeding birds from an Ecuadorian lowland forest. Bulletin of the British Ornithologists’ Club 124:28–37. (enlace)
- Greider, M., & E. S. Wagner (1960). Black Vulture extends breeding range northward. Wilson Bulletin 72:291. (enlace)
- Greiner, E. C., G. F. Bennett, E. M. White, & R. F. Coombs (1975). Distribution of avian hematozoa of North America. Canadian Journal of Zoology 53:1762–1787. (enlace)
- Griffiths, C. S. (1994). Monophyly of the Falconiformes based on syringeal morphology. Auk 111 (4):787-805. (enlace)
- Grube, G. E. (1953). Black Vulture breeding in Pennsylvania. Wilson Bulletin 65:119. (enlace)
- Grunberg, H. (1984). The autumn migration: Northwest Canada region. American Birds 38:223–224
- Haene, E. & Gill G. (1991). El Proyecto Parque Nacional Sierra de las Quijadas (Provincia de San Luis, República Argentina). Informe inédito APN. 100pp. (S.I.B.)
- Haene, E. (2007). Parque Nacional Sierra de las Quijadas. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 400-101. Temas de Naturaleza y Conservación 5. CD-ROM. Ed. Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires. (enlace)
- Hartert, E. & S. Venturi (1909). Notes sur les oiseaux de la République Argentine. Novit. Zool. 16: 159-267. (enlace)
- Hartman, F. A. (1955). Heart weight in birds. Condor 57:221–238. (enlace)
- Haverschmidt, F. (1947). The Black Vulture and the Caracara as vegetarians. Condor 49:210. (enlace)
- Haverschmidt, F. (1977). Allopreening in the Black Vulture. Auk 94:392. (enlace)
- Hayes, F. E. (1995). Status, Distribution and Biogeography of the Birds of Paraguay. Monographs in Field Ornithology, Nº1; 224 pp. (enlace)
- Hecker E. & F. Vidoz (1998). Aves del Lago Escondido. 95 pp. Hidden Lake S.A.
- Henckel, E. (1976). Turkey Vulture banding problem. North American Bird Bander 1:126. (enlace)
- Heredia, G. & M. Funes (1987). Informe técnico sobre el Parque provincial Tromen-Los Barros y Reserva Provincial Lagunas de Epu Lauquen. Subsecretaría de la Producción y Recursos Naturales de la Provincia del Neuquén. Infome Inédito.
- Heredia, J. (2006). Aves de las Sierras de Córdoba. Guía de campo. 50 p. La Falda. Córdoba
- Hervas, J. M., Gallo, Arias & F. Villafañe (2004). Informe Técnico. Registro ocasional de especies en el P.N. Talampaya Nº 2. 5 pp.
- Hill, J. R., III, & P. S. Neto (1991). Black Vultures nesting on skyscrapers in southern Brazil. Journal of Field Ornithology 62:173–176. (enlace)
- Hilty, S. L., Tudor, G. & Gwynne, J. A. (2003). Birds of Venezuela. (2º Ed.). Princeton University Press; 875 pp.
- Hudson, G. E. (1948). Studies on the muscles of the pelvic appendages in birds II: the heterogenous order Falconiformes. American Midland Naturalist 39:102-127. (enlace)
- Houston, D. C. (1985). Evolutionary ecology of Afrotropical and Neotropical vultures in forests. Ornithological Monographs 36:856–864. https://doi.org/10.2307/40168321. (enlace)
- Houston, D. C. (1986). Scavenging efficiency of Turkey Vultures in tropical forest. Condor 88:318–323. (enlace)
- Houston, D. C. (1988). Competition for food between Neotropical vultures in forest. Ibis 130:402–417. (enlace)
- Houston, D. C., & J. E. Cooper (1975). The digestive tract of the whiteback griffon vulture and its role in disease transmission among wild ungulates. Journal of Wildlife Diseases 11:306–313. (enlace)
- Hutton, J. & Schimff, S. (2001). Lista de Aves. Parque Nacional Mburucuyá (S.I.B.)
- Iñigo Elías, E. E. (1987). Feeding habits and ingestion of synthetic products in a Black Vulture population from Chiapas, Mexico. Acta Zoológica Mexicana 22:1–15. (enlace)
- Jackson, J. A. (1975). Regurgitative feeding of young Black Vultures in December. Auk 92:802–803. (enlace)
- Jackson, J. A., I. D. Prather, R. N. Conner & S. P. Gaby (1978). Fishing behavior of Black and Turkey Vultures. Wilson Bulletin 90:141–143. (enlace)
- Jackson, J. A. (1983). Nesting phenology, nest site selection, and reproductive success of Black and Turkey vultures. In Vulture Biology and Management (S. R. Wilbur and J. A. Jackson, Editors). University of California Press, Los Angeles, CA, USA; 245–270.
- Jones, H. L. & Vallely, A. C. (2001) Annotated Checklist of th Birds of Belize. Lynx Edicions. Barcelona; 71 pp.
- Juri, M. D. & J. M. Chani (2005). Variación en la composición de comunidades de aves a lo largo de un gradiente urbano (Tucumán, Argentina). Acta Zoológica Lilloana, 49 (1-2): 49-57. (enlace)
- Juri, M. D. (2007). Estudios ecológicos de la comunidad de aves en un gradiente urbano. Tesis doctoral, Facultad Ciencias Naturales e Instituto M. Lillo, Universidad Nacional de Tucumán, 276 pp. (enlace)
- Kiff, L. F., D. B. Peakall, M. L. Morrison, & S. R. Wilbur (1983). Eggshell thickness and DDE residue levels in vulture eggs. In Vulture Biology and Management (Wilbur & Jackson, Eds.). University of California Press, Los Angeles, CA, USA. pp. 440–458. (enlace)
- Kirk, D. A. (1988). Ecological separation of small Cathartid vultures in South America. Ph.D. dissertation, University of Glasgow, Glasgow, Scotland. (enlace)
- Kirk, D. A., & J. E. P. Currall (1994). Habitat associations of migrant and resident vultures in central Venezuela. Journal of Avian Biology 25:327–337. (enlace)
- Kirk, D. A., & A. G. Gosler (1994). Body condition varies with migration and competition in migrant and resident South American vultures. Auk 111:931–942. (enlace)
- Klimaitis, J. F. & F. N. Moschione (1986) Cartilla sistemática de las aves de la Reserva Integral de selva subtropical marginal de Punta Lara y sus alrededores (Provincia de Buenos Aires). (enlace)
- Klimaitis, J.F. & F. N. Moschione (1987) Aves de la Reserva Integral de Selva Marginal de Punta Lara y sus alrededores. Minist. Eco. Prov. Bs. Aires.
- Kovacs, C., O. Kovacs, Z. Kovacs & C. M. Kovacs (2005). Manual ilustrado de las aves de la patagonia, Antártica Argentina e islas del Atlántico Sur; 368 pp. El Bolsón
- Krabbe, N. (2000). Comparación entre ambientes con distintos tipos de intervención. P.N. Calilegua y alrededores. Inf. Inéd. APN-DRNO. (S.I.B.)
- Krauczuk, E. R., & Baldo, J. D. (2004). Contribuição para o conhecimento da avifauna de um fragmento de floresta com araucária em Misiones, Argentina. Atualidades Ornitológicas, 119(6). (enlace)
- Krauczuk, E. R. (2005). Aves do Inta-Campo Anexo ao Zaiman, e do Campus da Universidade Nacional de Misiones, Posadas, Misiones, Argentina. Atualidades Ornitológicas Nº 126: 1-22. (enlace)
- Lambertucci, S. A., Barbar, F., Cabrera, C., & Bertini, M. (2009). Comentarios sobre las aves de la Sierra de Pailemán, Río Negro, Argentina. Nuestras Aves, 54, 81-87. (enlace)
- Larochelle, J., J. Delson, & K. Schmidt-Nielsen (1982). Temperature regulation in the Black Vulture. Canadian Journal of Zoology 60:491–494. (abstract)
- Lemon, W. C. (1991). Foraging behavior of a guild of Neotropical vultures. Wilson Bull.. 103(4): 698–702. (enlace)
- Lepage, D. (2003). Avibase-La base de datos World Bird; Zopilote Negro (Coragyps atratus) (Bechstein, 1793). Recuperado el 19/01/2020 de https://avibase.bsc-eoc.org/species.jsp?avibaseid=4FF7DE80F4192276
- Ligon, J. D. (1967). Relationships of the cathartid vultures. Occas. Pap. Mus. Zool. Univ. Mich. 651. (enlace)
- Lindsay, D. S., D. S. Zarlenga, H. R. Gamble, F. Al-Yaman, P. C. Smith, & B. L. Blagburn (1995). Isolation and characterization of Trichinella pseudospiralis Garkavi, 1972 from a Black Vulture (Coragyps atratus). Journal of Parasitology 81:920–923. (enlace)
- López Lanús, B. (1997). Inventario de las aves del Parque Nacional “Río Pilcomayo” Formosa, Argentina. Monografía Especial L.O.L.A. Nº 4, 76 págs. Bs.As.
- López-Lanús, B., Grilli, P., Di Giácomo, A. S., Coconier, E., & Banchs, R. (2008). Categorización de las Aves de la Argentina, según su estado de conservación. Informe de Aves Argentinas/AOP y la Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina.
- López-Lanús, B., A. Galimberti, C. Giarduz, H. Luna, M. Manassero, A. A. Pautasso, M. P. Ducommun, A. A. G. Saigo, P. Petracci, G. A. Marteleur & M. R. de la Peña (2013a). Inventario focal de fauna de las estancias El Estero, La Norma, El Matrero y La Elena en el sitio piloto San Javier: Santa Fe, Argentina. En pp. 135-169/277-306/331-347/507,511-513: G. D. Marino, F. Miñarro, M. E. Zaccagnini y B. López-Lanús (eds.). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. (enlace)
- López-Lanús, B., A. S. Di Giacomo, A. Azpiroz, P. Haynes, A. Galimberti, A. Keyel, A. Ocampo, R. Güller, R. Moller Jensen, M. Mattalia, H. Cardoso, C. Giarduz, G. Papini & A. G. Di Giacomo (2013b). Inventario focal de fauna de las estancias La Higuera, María Concepción, La Serena y Virocay en el sitio piloto Aguapey: Corrientes, Argentina. En pp. 179-223/277-293/307-319/331-347/490,494-506: G. D. Marino, F. Miñarro, M. E. Zaccagnini y B. López-Lanús (eds.). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas Nº 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. (enlace)
- López-Lanús, B. & P. Mencia (2017). Diversidad, abundancia y estado de conservación de las aves de los riachos Monte Lindo y Pilagá en el este de la provincia de Formosa, Argentina. Nótulas Faunísticas 228: 1-29. (enlace)
- Lovell, H. B. (1947). Black Vultures kill young pigs in Kentucky. Auk 64:131–132. (enlace)
- Lucero, M. M. (1983). Lista y distribución de aves y mamíferos de la provincia de Tucumán. Miscelánea No 75. Fund. M. Lillo. Tucumán. Págs. 61.
- Lucero, M. M., J. M. Chani, Z. J. Brandán, A. L. Echevarría & M. D. Juri (2002). Lista de aves de San Miguel de Tucumán y Yerba Buena. Acta Zool. Lilloana 46 (1 y 2): 131-135. (enlace)
- Lucero, M. M., Z. J. Brandán & J. M. Chani (2005). Composición y variación anual de la avifauna de los tres grandes parques urbanos de San Miguel de Tucumán (Tucumán, Argentina). Acta Zool. Lilloana 49 (1 y 2): 43-48.
- Lucero, F. (2013). Listado de las aves observadas en la Reserva Natural Municipal Articulada: Laguna Guanacache, Laguna del Toro y Bañados del Carau, provincia de San Juan y Laguna Seca y los Bañados del Tulumaya, provincia de Mendoza, Argentina. Nótulas Faunísticas (segunda serie), 137: 1-16. (enlace)
- Lucero, F. (2020). Primeras evidencias para especies y subespecies de aves en la Provincia de San Juan, Argentina. EcoRegistros Revista, 10 (8): 38-52. (enlace)
- Llanos, F., Failla, M., García. G., Giovine, P., Carbajal, M., Gonzaléz, P., Paz Barreto, D., Quillfeldt, P. & Masello, J. (2011). Birds from the endangered Monte, the Steppes and Coastal biomes of the province of Río Negro, northern Patagonia, Argentina. Check List and Authors. Journal of species list and distribution. (enlace)
- Maceda, J. J., M. Mero, H. Riesco & M. Dolsan (2001). Lista de las aves de la reserva Parque Luro, La Pampa. Rev. Fac. Agr. UNL La Pampa. Vol. 12 Nº 2: 47-59. (enlace)
- Mactavish, B. (1993). The autumn migration: August 1 to November 30-1992, Atlantic Provinces region. American Birds 47:55–59. (enlace)
- Malizia, L. R., P. G. Blendinger, M. Eva Alvarez, L. O. Rivera, N. Politi & G. Nicolossi (2005). Bird communities in Andean premontane forests of Northwestern Argentina. Ornitología Neotropical 16: 231-251. (enlace)
- Mamaní, J. C., O. B. Quiroga, S. Aveldaño & T. G. Moreno Ten (2018). La avifauna de Yavi, provincia de Jujuy, Argentina. Nótulas Faunísticas. Segunda Serie 245: 1-19. (enlace)
- Marateo, G., Povedano, H., & Alonso, J. (2009). Inventario de las aves del Parque Nacional El Palmar, Argentina. Cotinga, 31(1), 47-60. (enlace)
- Marchant, S. (1960). The breeding of some S.W. Ecuadorian birds. Ibis 102(2):349–382. (enlace)
- Marelli, C.A. (1933) Aves observadas en el sur de la provincia de Buenos Aires. Hornero 5: 193-199. (enlace)
- Marigliano, N. L., C. M. Antelo, Z. J. Brandán Fernández & C. I. Navarro (2017). Avifauna asociada a sectores de bosques ribereños con modificaciones antropogénicas (Tucumán, Argentina). III. Composición y cambios estacionales del ensamble. Acta Zoológica Lilloana. 61 (1): 15-26. (enlace)
- Marone, L. (1992). Estatus de residencia y categorización trófica de las especies de aves en la Reserva de la Biosfera de Ñacuñan, Mendoza. El Hornero 13: 207-210. (enlace)
- Márquez, J., Chébez, J. C., Flores, A., Haene, E. & Sánchez, E. (1991) Sistema de Áreas Naturales Protegidas de la Provincia de San Juan (República Argentina). (enlace)
- Márquez, J. (1999). Las áreas protegidas de la provincia de San Juan. Multequina, (8), 1-10. (enlace)
- Martínez Gamba, R. (2014). Lista de aves del Parque Natural Municipal Monte Seguín, Provincia de Misiones, Argentina. Nótulas Faunísticas. Segunda Serie. 163: 1-10 (enlace)
- Masramón, D. O. de (1983). Lista de aves del nordeste de San Luis. Hornero extraordinario 77-87. (enlace)
- Matarasso, H. F. (2007). Checklist. Aves del Neuquén. (enlace)
- McHargue, L. A. (1977). Nesting of Turkey and Black Vultures in Panama. Wilson Bulletin 89:328–329. (enlace)
- McHargue, L.A. (1981). Black Vulture nesting, behaviour and growth. Auk. 98(1): 183–185. (enlace)
- McIlhenny, E. A. (1939). Feeding habits of Black Vulture. Auk 56:472–474. (enlace)
- McIlhenny, E. A. (1945). An unusual feeding habit of the Black Vulture. Auk 62:136–137. (enlace)
- Melo, M. A., A. F. Batisteli, R. Guillermo-Ferreira, & A. J. Piratelli (2018). “Wanted dead or alive:” Black Vultures (Coragyps atratus) feeding on a live Capybara (Hydrochoerus hydrochaeris). Ornitología Neotropical 29:91–93. (enlace)
- Mérida, E. & A. Bodrati (2006). Consideraciones sobre la conservación de los talares de barrancas del nordeste de Buenos Aires y descripción de las características de un relicto en Baradero. Pp.: 71-82, en: Mérida, E. & J.Athor (Editores). Talares bonaerenses y su conservación. Fundación de Historia Natural “Félix de Azara”. Buenos Aires. (enlace)
- Miatello, R., J. Baldo, M. Ordano, C. Rosacher & L. Biancucci (1999). Avifauna del Parque Nacional Quebrada del Condorito y Reserva Hídrica Provincial de Achala. Córdoba. Argentina. Secretaría de Agricultura, Ganadería y Recursos Renovables. Córdoba; 193 pp.
- Miatello, R. & Fernandez, F. (2015). Informe de comisión a la Reserva de la Defensa La Calera y relevamiento de Fauna Silvestre (S.I.B.)
- Monroe, B. L., Jr. (1968). A Distributional Survey of the Birds of Honduras. Ornithological Monographs 7. American Ornithologists’ Union, Washington, DC, USA. 458 pp. (enlace)
- Morici, A. (2012). Las aves del Área Natural Protegida “Dique Roggero” (Buenos Aires, Argentina). Biológica 15; 24-36. (enlace)
- Moschione, F. & L. Bishels (2005). Listado de las aves del Parque Provincial Loro Hablador, provincia del Chaco. Informe Técnico. Proyecto Elé/DFS. Secretaría de Ambiente y Desarrollo Sustentable de la Nación
- Moschione, F. N. & Segovia, J. (2005). Listado de las aves de la Reserva Natural Las Lancitas, Santa Bárbara, Jujuy. Informe técnico Proyecto Elé/DFS, Secretaría de Ambiente y Desarrollo Sustentable de la Nación.
- Moschione, F.N. & M. A. González (2007). Aves de la Reserva Natural General Pizarro. Departamento Anta, Provincia e Salta. Lista preliminar de la RN en formación, actualizada a marzo de 2007. Inf. Inéd. APN-DRNO.12 pp. (S.I.B.)
- Moschione, F., Spitznagel, O. & M. Gonzalez (2014). Lista de Aves de Salta (Birds Checklist). Ministerio de Cultura y Turismo de Salta. Argentina. 47 p. (enlace)
- Mouchard, A. (2017). Interacción agonística de un Águila mora (Geranoaetus melanoleucus) seguida de consumo de un Jote cabeza negra (Coragyps atratus). Nuestras Aves 62:31-32. (enlace)
- Mrosovsky, N. (1971). Black Vultures attack live turtle hatchlings. Auk 88:672–673. (enlace)
- Narosky, T. & L. M. Lozzia (1988). Aves de la Reserva El Bagual. Nuestras Aves 15: 7-9. (enlace)
- Narosky, T. & A. G. Di Giacomo (1993) Las aves de la provincia de Buenos Aires, distribución y estatus. Asoc. Orn. del Plata. Vazquez Mazzini Editores, L.O.L.A., Buenos Aires
- Narosky, T., & Chebez, J. C. (2002) Guía para la identificación de las aves de Iguazú. Buenos Aires: Vázquez Mazzini. 128 pp.
- Narosky, T. & D. Yzurieta (2010). Aves de Argentina y Uruguay: guía de identificación. (16ª Ed.). Vázquez Mazzini Editores, Argentina; 432 pp.
- Navarro, C. I., Z. J. Brandán, C. M. Antelo & N. L. Marigliano (2000). Avifauna invernal en una localidad del Bosque Chaqueño Serrano (Las Tipas, Tucumán, Argentina). Acta Zool. Lilloana 45 (2): 233-240.
- Navarro, C. I. & Marigliano, N. L. (2006). Avifauna otoño-invernal en un sector de bosque Chaqueño Serrano (Trancas, Tucumán, Argentina). Acta Zoológica Lilloana, 50 (1-2): 55-60. (enlace)
- Navarro, C., Brandán, Z. J. & Marigliano, N. L. (2007). Lista comentada de las aves de un fragmento de la Reserva Provincial La Florida, Tucumán. Acta Zoológica Lilloana, 51 (2): 142-150. (enlace)
- Navas, J. R. (1982). Introducción a la avifauna del Parque Nacional El Palmar. Anales P. Nac. XV: 35-64
- Nellar Ramonella, M. M. (1993). Aves de la provincia de San Luis. Lista y distribución. Ed. Mus. Priv. Cienc. Nat. e Inv. Ornit. “G. E. Hudson”. San Luis. Págs. 98.
- Ng, D., & B. D. Jasperson (1984). Interspecific allopreening between Crested Caracara and Black Vultures. Condor 86:214–215. (enlace)
- Nores, M., D. Yzurieta & R. Miatello (1983). Lista y distribución de las aves de Córdoba, Argentina. Boletín de la Academia Nacional de Ciencias de Córdoba. 56: 1-96.
- Nores, M., D. Yzurieta & S. A. Salvador (1991). Lista y distribución de las aves de Santiago del Estero, Argentina. Bol. Acad. Nac. Cs.Córdoba, 59: 157-196. (enlace)
- Olivera, D. (2008-2011). Relevamiento de Aves Reserva Senderos del Monte de la Familia Powell, Departamento de Gualeguaychú, Provincia de Entre Ríos Argentina. avesargentinas.org.ar (enlace)
- Olivera, L. R. (2015) Reflejos en las aguas del Moconá. Naturaleza e Historia del Parque Provincial Moconá, su entorno y su gente. (1º Ed.) C.A.B.A. Fundación de Historia Natural Félix de Azara. 184 pp.
- Olrog, C. C. (1959) Las Aves Argentinas. Una Guía de campo. Universidad Nacional de Tucumán. Instituto “Miguel Lillo”. Tucumán. Argentina. 343 pp.
- Olrog, C. C. (1979). Nueva Lista de la Avifauna Argentina. Ministerio de Cultura y Educación-Fundación Miguel Lillo Tucumán, Argentina. Opera Lilloana, XXVII:58 (enlace)
- Olson, S. L., H. Loftin, & J. Wiese (1967). Observations on the behavior of Black and Turkey Vultures at traps and in captivity. Bird-Banding 38:75–76. (enlace)
- Ordano, M. (1993). Composicion y Estructura de una Comunidad de Aves Altoserranas (Cordoba), entre 1800 y 1900m Durante un Ciclo Anual. Tesina de Grado. Catedra de Zoologia II, FCEF y N, UNC. 52Pp (S.I.B.)
- Ordano, M. (1996). Estudio de una comunidad de aves altoserrana (Córdoba, Argentina) durante un ciclo anual. Rev. Asc. Cienc. Nat. Litoral 27 (2): 83-94. (enlace)
- Ordano, M., A. Bortoluzzi, M. Chatellenaz & L. Biancucci (2008). Respuesta a corto plazo de las aves a la quema y al pastoreo en pastizales del Parque Nacional Mburucuyá, Argentina. INSUGEO, Miscelánea, 17 (2): 425-446. (enlace)
- Ortiz, G. & F. Murúa (1994). Aves de ambientes acuáticos de la provincia de San Juan (Argentina). I. Multiquina, 3: 125-131. (enlace)
- Ortiz, D. & P. Capllonch (2007). Guía de los rapaces de la provincia de Tucumán. Tucumán, República Argentina. (enlace)
- Ortiz, D. & C. Ruiz (2011a). Migración y estacionalidad de aves en el arroyo Mista, Tucumán, Argentina. Nuestras Aves 56: 16-18. (enlace)
- Ortiz, D., R. Aráoz & C. Aldetere (2011b). Lista sistemática de las aves de la reserva experimental Horco molle, provincia de Tucumán, Argentina. Nótulas Faunísticas. Segunda Serie. No 62: 1-5. (enlace)
- Ortiz, D. & R. Aráoz (2014). Aves de la Sierra de San Javier. Universidad Nacional de Tucumán.
- Ortiz, S. G., H. A. Gómez, R. V. Blanco Fager, L. Videla & E. Godoy Luna (2016a). Aves de las diferentes ecorregiones, San Juan, Argentina. En: Martínez Carretero E. (ed.). San Juan Ambiental. Pp. 494
- Ortiz, S. G., Fager, R. V. B., & Gómez, H. A. (2016b). San Juan, Argentina. Eduardo Martínez Carretero, 259. (enlace)
- Ortiz, D., E. Martínez Paster, J. P. Juliá & P. Quiroga (2018a). Lista de aves de la Reserva Privada San Pablo, provincia de Tucumán, Argentina. Nótulas Faunísticas. Segunda Serie 246: 1-8. (enlace)
- Ortiz, S. G., V. Blanco Fager, M. G. Cánovas, H. J. Villavicencio, L. Videla, A. Laspina & P. Asunto (2018b). Listado de las aves urbanas y periurbanas con límite rural en la ciudad de San Juan, Argentina. Nótulas Faunísticas 254: 1-7. (enlace)
- Pagano, L. G. & E. Mérida (2009) Aves del Parque Costero del Sur. Pp. 200–244, en: Athor J. (de) Parque Costero del Sur. Naturaleza, conservación y patrimonio cultural. Fundación de Historia Natural Félix de Azara, Buenos Aires, Argentina. (enlace)
- Pagano, L. G., E. A. Jordan, J. I. Areta, R.F. Jensen & I. Roesler (2012). Aves de la Reserva Natural Punta Lara. En 97-143: Roesler, I. y M.G. Agostini (eds). Inventario de los Vertebrados de la Reserva Natural Punta Lara, provincia de Buenos Aires, Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas Nº 8. Buenos Aires, Argentina. (enlace)
- Pagano, L. G., & S. A. Salvador (2017). Datos de pesos de aves argentinas. Parte 4. Historia Natural 7:21–43. (enlace)
- Pardieck, K. L., D. J. Ziolkowski Jr., M. Lutmerding, V. I. Aponte, & M-A. R. Hudson (2020). North American Breeding Bird Survey Dataset 1966–2019: U.S. Geological Survey data release. https://doi.org/10.5066/P9J6QUF6 (enlace)
- Parker, P. G., T. A. Waite, & M. D. Decker (1995). Kinship and association in communally roosting Black Vultures. Animal Behaviour 49:395–401. (abstract)
- Parker III, T. A., & Goerck, J. M. (1997). The importance of national parks and biological reserves to bird conservation in the Atlantic forest region of Brazil. Ornithological Monographs, 527-541. (enlace)
- Parmalee, P. W. (1954). The vultures, their movements, exonomic status and control in Texas. Auk. 71(4): 443–453. (enlace)
- Parmalee, P. W. & Parmalee, B. (1967). Results of banding studies of Black Vultures in eastern North America. Condor. 69(2): 146–155. (enlace)
- Parrott, G. C. (1970). Aerodynamics of gliding flight of a Black Vulture Coragyps atratus. Journal of Experimental Biology 53:363–374. (enlace)
- Partridge, W. H. (1953). Observaciones sobre aves de las provincias de Córdoba y San Luis. Hornero 10: 23-73. (enlace)
- Pastore, H. (1994). Lista de aves observadas en el Parque Nacional Los Alerces, 1994. (S.I.B.)
- Paz Barreto, D. (1990). Lista sistemática de aves observadas en la reserva de fauna de punta Bermeja. Prov. de Rio Negro. Dirección de Planificación Ambiental, Provincia de Rio Negro. Banco de datos AOP: PRION 218
- Paz Barreto, D. (1996). Aves de la provincia de Rio Negro. Lista y bibliografía. Consejo de Ecología y Medio Ambiente, deleg. S.C. de Bariloche. (S.I.B.)
- Paz Barreto, D. (1997). Aves de la Provincia de Río Negro. II Localidades confirmadas. Consejo de Ecología y Medio Ambiente, Delegación Bariloche. Pp. 1-24. (enlace)
- Paz Barreto D. (2018). Registro de Aves del Parque Nacional Laguna Blanca de 1951 a 2018. Informe Técnico Parque Nacional Laguna Blanca, Administración de Parques Nacionales, Zapala, Neuquén. 25 pp. (enlace)
- Pennycuick C. J. (1973). The soaring flight of vultures. In Scientific American: 38-45. (enlace)
- Pennycuick, C. J. (1983). Thermal soaring compared in three dissimilar tropical bird species, Fregata magnificens, Pelecanus occidentalis and Coragyps atratus. Journal of Experimental Biology 102:307–325. (enlace)
- Perellano, M. & Basti, A. (1981). Lista de fauna observada en la Seccional Perito Moreno, Parque Nacional Nahuel Huapi. (S.I.B.)
- Pereyra, J. A. (1927). Notas ornitológicas. Pichón de Cuervo (Cathartes atratus brasiliensis). Hornero 4: 80. (enlace)
- Pereyra, J. A. (1937a). Aves nuevas para la provincia de Buenos Aires. Hornero 6: 438-439. (enlace)
- Pereyra, J. A. (1937b). Contribución al estudio y observaciones ornitológicas de la zona norte de la Gobernación de La Pampa. Mem. Jard. Zool. La Plata. Tomo 7: 197-326
- Pereyra, J. A. (1945). Las aves del territorio del Neuquén. An. Mus. de la Pat. Perito “Francisco P. Moreno”, Adm. Prq. Nac., Argentina. T. I, pags. 61-99. Lam. I-VIII
- Pereyra, J. A. (1950a). Avifauna argentina. Hornero 009 (02): 178-241. (enlace)
- Pereyra, J. A. (1950b). Las aves del territorio de Misiones. Anal. Mus. Nahuel Huapi Pto. F. P. Moreno 1-40. Buenos Aires
- Pereira, J. & N. Fracassi (1999). Registro de observación de especies. Parque Nacional El Rey. Inf. inéd. APN-DRNO. 4 pp. (S.I.B.)
- Pérez, C. H. F. & P. Petracci (1998). Notas sobre la avifauna de la cuenca media y superior del Río Negro, Provincia de Río Negro, Argentina. X RAO: 38.
- Perovic, P.G., De Bustos, S., Tálamo, A., Trucco, C. & D. Vázquez (2000). Informe Final del Relevamiento de Flora y Fauna en el Parque Nacional Calilegua y alrededores, Jujuy, Argentina. Monitoring and Modelling the Impacts of Changing Governments Policies on Biodiversity Conservation in the Andes. Proyecto Bioandes. First Annual Report.Unión Europea. 34 pp. (S.I.B.)
- Petracci, P. F., K. J. V. Delhey, C. H. F. Pérez, D. Basanta, M. V. Massola, M. Zamorano & M. Fosatti (2004). Nuevos aportes al conocimiento de la distribución y anidación de algunas especies de aves en la Argentina. Nuestras Aves 48: 25-31. (enlace)
- Piacentini, H., D. Paz Barreto, & A. Wucusich (2003). Registro de Fauna zona de Reserva Cinco Chañares págs. 369-386, en Bajo del Gualicho, una planicie patagónica bajo el nivel del mar-Realidad y leyenda. Coord. R. F. Massera y J. C. Guarido. Secretaría de Estado de Acción Social de Río Negro 2003, págs. 1-416. (enlace)
- Planqué, R., Vellinga, W.P., Pieterse, S., Jongsma J. & de By, R. (Xeno-canto Foundation). Zopilote negro (Coragyps atratus) (Bechstein, 1793); Recuperado de https://www.xeno-canto.org/species/Coragyps-atratus, el 19/01/2020
- Prather, I. D., R. N. Conner, & C. S. Adkisson (1976). Unusually large vulture roost in Virginia. Wilson Bulletin 88:667–668. (enlace)
- Presti, P. M. & A. L. Echevarría (2009). El ensamble de aves en un relicto de selva pedemontana: Parque Percy Hill (Yerba Buena, Tucumán, Argentina). Acta Zoológica Lilloana, 53 (1-2): 64-76. (enlace)
- Prins, T. G., Reuter, J. H., Debrot, A. O., Wattel, J. & Nijman, V. (2009). Checklist of the birds of Aruba, Curaçao and Bonaire, south Caribbean. Ardea. 97(2): 137–268. (enlace)
- Pugnali, G. & P. Chamorro (2008). Lista de aves de la Reserva Costanera Sur. Aves Argentinas. Buenos Aires. (enlace)
- Pujalte, J. C., A. R. Reca, A. Balabusic, P. Canevari, L. Cusato & V. P. Fleming (1995). Unidades ecológicas del Parque Nacional Río Pilcomayo. Anales de Parques Nacionales 16: 1-185. (enlace)
- Pütz, C. C., Basso, M., de los Llanos, E. R., & Kufner, M. (2012). La fauna chaqueña de Córdoba (Argentina) afectada por la transformacion agricola. Ecología Aplicada, 11(1-2), 77-87. (enlace)
- Quiroga, O. B., Aveldaño, W. S., Mamaní, J. C., Martínez-Pastur, E. & Moreno-Ten, Th. (2020). Notas sobre veintidos especies de aves del noroeste argentino. Nótulas Faunísticas- Segunda Serie, 285:1-7. (enlace)
- Rabenold, P. P. (1986). Family associations in communally roosting Black Vultures. Auk 103(1): 32–41. (enlace)
- Rabenold, P. P. (1987a). Recruitment to food in Black Vultures: evidence for following from communal roosts. Animal Behaviour 35: 1775–1785. (abtract)
- Rabenold, P. P. (1987b). Roost attendance and aggression in Black Vultures. Auk 104(3): 647–653. (enlace)
- Raffaele, H., Wiley, J., Garrido, O., Keith, A. & Raffaele, J. (2003). Birds of the West Indies. Published by Princeton University Press 41 William Street Princeton, New Jersey 08540; 216 pp.
- Ramilo, E. J., Chehébar, C. E. & Mazzucchelli, S. (1993). Plan General de Manejo. Parque Nacional Laguna Blanca. APN. (enlace)
- Rea, A. M. (1983). Cathartid affinities: a brief overview. In Vulture Biology and Management (S. R. Wilbur and J. A. Jackson, Editors). University of California Press, Los Angeles, CA, USA. pp. 26–54.
- Regidor, H.A., Acuña, E., Orce, H., Sosa, A. & S. Castrillo (2005). Relevamiento de la Biodiversidad en Pizarro, Departamento de Anta, Salta, y los efectos de los desmontes sobre la misma. IRNED – Instituto de Recursos Naturales y Ecodesarrollo. Universidad Nacional de Salta. 39 pp. (S.I.B.)
- Richardson, D. M. (1989). Close nesting of a Black Vulture and a Turkey Vulture. Wilson Bulletin 101:639–640. (enlace)
- Ricklefs, R. E. (1973). Patterns of growth in birds: II. Growth rate and mode of development. Ibis 115:177–201. DOI: 10.1111/j.1474-919X.1973.tb02636.x
- Ridgely, R.S. & Gwynne Jr., J.A. (1989). A Guide to the Birds of Panamá, with Costa Rica, Nicaragua, and Honduras (2º Ed.) Princeton University Press. 534 pp.
- Roads, K. M. (1936). Black Vultures kill and eat new-born lambs. Wilson Bulletin 48:219. (enlace)
- Robbins, M. B., & D. A. Easterla (1992). Birds of Missouri: Their Distribution and Abundance. University of Missouri Press, Columbia, MO, USA. Pp: 61-62
- Roda, M.A. & C.F. Danti (2015). Nuevos registros de aves para Saladillo, provincia de Buenos Aires. Nuestras Aves 60:32-38. (enlace)
- Rodríguez Graves, V., A. Pautasso, P. Preliasco, M. V. Del Sastre & F. Miñarro (2009). Relevamiento ambiental de la Estancia Isleta Linda. Provincia de Santa Fe. Fundación Vida Silvestre Argentina.
- Rodríguez, E. D. (2012). Aves del Cerro San Bernardo y de las Serranías del Este de la Ciudad de Salta. (1º Ed.) Salta: Fondo Editorial Secretaría de Cultura de la Provincia de Salta.
- Roesler, I. (2001). Nuevas citas de aves para al partido de General Villegas, provincia de Buenos Aires, Argentina. Nuestras Aves 41: 32-33. (enlace)
- Roesler, I. & Agostini, M.G. (2012). Inventario de los vertebrados de la Reserva Natural Punta Lara, provincia de Buenos Aires, Argentina. (enlace)
- Roig, V. G. (1965). Elenco sistemático de los mamíferos y aves de la provincia de Mendoza y notas sobre su distribución geográfica. Bol. de Estud. Geogr. 12 (49): 175-222. (enlace)
- Romiti, D. (1997). Avistajes de fauna. Parque Nacional Laguna Blanca. 5 pág. Nota 7, DRP, 3/04/1997. (S.I.B.)
- Rosso, P. (2002). Listado de aves observadas en la jurisdicción de la Seccional Río Arrayanes. Parque Nacional Los Alerces. (S.I.B.)
- Saibene, C.A., Castelino, M.A., Rey, N.R., Herrera, J. & Calo, J. (1996). Inventario de las aves del Parque Nacional “Iguazú”, Misiones, Argentina. Monografía, (9); 68 pp.
- Salvador, S. A. (1990). Nidificación de rapaces argentinos (Falconiformes y Strigiformes). Nuestras aves 23: 28-29. (enlace)
- Salvador, S. A. (2014a). Peso de las aves del Departamento General San Martín, Córdoba, Argentina. Biológica 17: 48-57. (enlace)
- Salvador, S. A. & Salvador, L. A. (2014b). Aves de Villa María y del departamento General San Martín, Córdoba, Argentina. Xolmis, 36, 1-18. (enlace)
- Salvador, S. A. (2016a). Distribución reproductiva de las aves de Argentina y sus territorios. Recopilación inédita. Córdoba, Argentina. (enlace)
- Salvador, S. A. (2016b). Registros de depredadores de huevos, pichones y volantones de aves de Argentina. Acta Zoológica Lilloana 60. (enlace)
- Salvador, S. A., L. A. Salvador & C. Ferrari (2017). Aves de la provincia de Córdoba. Distribución e Historia Natural. Ed. DP. Argentina. SA.
- Sanzín, R. (1918). Lista de aves mendocinas. Hornero 1: 147-152. (enlace)
- Sarquis, J. A. (2018). Conservación de la avifauna de Entre Ríos (Argentina): uso de métodos biogeográficos y de optimización para evaluar la efectividad de las áreas protegidas. Tesis. Laboratorio de Biodiversidad y Conservación de tetrápodos, INALI-CONICET-UNL. (enlace)
- Saunders, A. A. (1906). The habits of the Black Vulture in Nicaragua. Bird-Lore 8:165–167. (enlace)
- Sazima, I. (2011). Black Vulture (Coragyps atratus): bath and drink. Revista Brasileira de Ornitologia 19:81–84. (enlace)
- Scherer-Neto, P. & Costa-Straube, F. (1995). Aves do Paraná: (história, lista anotada e bibliografia). Curitiba-Paraná: Ed. dos autores, Brasil; 117 pp. (enlace)
- Schulenberg, T. S., Stotz, D. F., Lane, D. F., O’Neill, J. P., Parker III, T. A. & A. Brack Egg (2010). Birds of Perú. Revised and Updated Edition ed. Princeton University Press; 664 pp.
- Segura, L. N., N. Bogado, C. A. Darrieu & D. Montalti (2014). La colección de huevos “Pablo Girard” del Museo de La Plata. Probiotica. FCN y M, UNLP, La Plata, Argentina, Serie Técnica y Didáctica 27: 1-14. (enlace)
- Seibold, I. & A. J. Helbig (1995). Evolutionary history of New and Old World vultures inferred from nucleotide sequences of the mitochondrial cytochrome b gene. Philosophical Transactions of the Royal Society of London B 350:163–178. https://doi.org/10.1098/rstb.1995.0150
- Serié, P. & C. H. Smyth (1923). Notas sobre aves de Santa Elena (E. Ríos). El Hornero. 003(01):037-055. (enlace)
- Serrano, G., Moreira, M., Natale, E., Monguillot, J. & R. Neira (2003). Informe Técnico. Registro ocacional de especies del Parque Nacional Sierra de Las Quijadas Nº 5. 3 pp.
- Sferco, G. D. & Nores, M. (2003). Lista comentada de las aves de la Reserva Natural Chancaní, Córdoba, Argentina. El Hornero, 18(1), 21-29. (enlace)
- Sibley, C. G. & J. E. Ahlquist (1990). Phylogeny and Classification of Birds: A Study in Molecular Evolution. Yale University Press, New Haven, CT, USA; 976 pp. https://doi.org/10.2307/j.ctt1xp3v3r
- Siegenthaler, G., E. Fiorucci, S. Tiranti, P. Borraz, M. Urioste & A. García (1990). Informe de avance del plan de relevamiento de los vertebrados de la provincia de La Pampa. Agro Pampeano. Ministerio de Asuntos Agrarios de La Pampa. 18: 38-49.
- Siegenthaler, G. B. (Coord.) (2004). Relevamiento de los vertebrados de la provincia de La Pampa. Inventario integrado de los recursos naturales de la provincia de La Pampa. Clima, geomorfología, suelo, vegetación y fauna de vertebrados. Reedición. INTA.UNLP. Gobierno de La Pampa, CD, multimedia. (enlace)
- Sick, H. (1997). Ornitologia Brasileira. Uma Introdução. Universidad de Brasília, Brasília, Brazil.
- Silva, C. & R. S. Carmo (2015). Comportamento allopreening entre urubu-de-cabeça-preta (Coragyps atratus) e caracará (Caracara plancus) no nordeste Brasileiro. Atualidades Ornitológicas 184:22. (enlace)
- Skutch, A. F. (1969). Notes on the possible migration and the nesting of the Black Vulture in Central America. Auk 86:726–731 (enlace)
- Slud, P. (1964). The birds of Costa Rica: distribution and ecology. Bulletin of the American Museum of Natural History 128:1–430. (enlace)
- Smyth, C. H. (1927). Descripción de una colección de huevos de aves argentinas. El Hornero. 004(01):001-016. (enlace)
- Sosa, H. J. (1995). Actualización de la lista de avifauna de la Reserva Provincial Laguna Llancanelo, Malargüe, Mendoza. Presencia estacional, preferencia de hábitats y nidificación. Multequina, (4), 65-75. (enlace)
- Sosa, H. J. (1999). Descripción del evento reproductivo del Flamenco austral (Phoenicopterus chilensis) en Laguna Llancanelo, Malargüe, Mendoza. Multequina 8: 87-99. (enlace)
- Sosa, M. & Mutt Coll, M. S. (2001). Aves de San Martín de los Andes y sus alrededores. Planilla de registro. 2001. (S.I.B.)
- Souto, H. N., A. G. Franchin, & O. M. Júnior (2009). New record of allopreening between Black Vultures (Coragyps atratus) and Crested Caracara (Caracara plancus). Sociobiology 53:1–5. (enlace)
- Sprunt, A., Jr. (1946). Predation on living prey by the Black Vulture. Auk 63:260–261. (enlace)
- Stager, K. E. (1964). The role of olfaction in food location by the Turkey Vulture (Cathartes aura). Los Angeles County Museum Contributions in Science 81:1–63. (enlace)
- Stempelmann, H. & F. Schulz (1890). Enumeración de las aves de la provincia de Córdoba. (República Argentina). Bol. Acad. Nac. Cienc. Córdoba 10: 393-409
- Steullet, A. B. & E. A. Deautier (1935). Catálogo sistemático de las aves de la República Argentina. Obra del Centenario del Museo de La Plata. Unv. Nac. de La Plata. Buenos Aires: Imprenta y Casa Editora «Coni». Tomo 1 (1º entrega): 1-256.
- Steullet, A. B. & Deautier, E. A. (1936). Catálogo sistemático de las aves de la República Argentina. Obra del Cincuentenario del Museo de La Plata. Unv. Nac. de La Plata. Buenos Aires: Imprenta y Casa Editora «Coni». Tomo 1 (1º entrega): 1: 257- 492
- Stewart, P. A. (1974). A nesting of Black Vultures. Auk 91(3): 595–600. (enlace)
- Stewart, P. A. (1978). Behavioral interactions and niche separation in Black and Turkey Vultures. Living Bird 17: 79–84. (enlace)
- Stewart, P. A. (1983). Adoption of introduced young and neglect of own by nesting Black Vultures. Wilson Bulletin 95:310–311. (enlace)
- Stewart, P. A. (1984). Population decline of Black Vultures in North Carolina. Chat 48:65–68. (enlace)
- Stiles, F. G., & A. F. Skutch (1989). A Guide to the Birds of Costa Rica. Christopher Helm, London, UK.
- Stolen, E. D. (1996a). Black and Turkey Vulture interactions with Bald Eagles in Florida. Florida Field Naturalist 24:43–45. (enlace)
- Stolen, E. D. (1996b). Roosting behavior and foraging ecology of Black Vultures in central Florida. M.S. thesis, University of Central Florida, Orlando, FL, USA.
- Stolen, E. D. (2000). Foraging behavior of vultures in central Florida. Florida Field Naturalist, 28(4), 173-181. (enlace)
- Stolen, E. D. & Taylor, W. K. (2003). Movements of black vultures between communal roosts in Florida. The Wilson Journal of Ornithology, 115(3), 316-320. (enlace)
- Stotz, D. F., Fitzpatrick, J. W., Parker III, T. A. & Moskovits, D. K. (1996). Neotropical birds: ecology and conservation. University of Chicago Press.
- Stresemann, E. & D. Amadon (1979). Order Falconiformes. In Check-list of birds of the world (Mayr & Cottrell, Eds.). Vol:1. Museum of Comparative Zoology, Cambridge, MA, USA. pp. 271–425. (enlace)
- Sweeney, T. M. (1984). Black and Turkey Vulture roost dynamics, marking, morphology and nesting in Virginia. Master’s Thesis, Virginia Polytechnic Inst. and State Univ., Blacksburg, VA. (enlace)
- Sweeney, T. M., J. D. Fraser & J. S. Coleman (1985). Further evaluation of marking methods for Black and Turkey Vultures. Journal of Field Ornithology 56:251–257. (enlace)
- Sweeney, T. M. & J. D. Fraser (1986). Vulture roost dynamics and monitoring techniques in southwest Virginia. Wildlife Society Bulletin 14:49–54. (enlace)
- Tagliarini, M. M., C. Y. Nagamachi, J. C. Pieczarka & E. H. C. de Oliveira (2007). Description of two new karyotypes and cytotaxonomic considerations on Falconiformes. Revista Brasileira de Ornitologia 15:261–266. (enlace)
- Tate, Jr., J. (1986). The blue list for 1986. American Birds 40:227-236. (enlace)
- Tejerina, P., M. J. Turnes & S. L. Tiranti (2006). Aves. En: Salomone, F. y N. Gouts. (eds.). Los vertebrados de las áreas protegidas de la provincia de La Pampa. Ed. Santa Rosa, La Pampa, Argentina. (enlace)
- Teulings, R. P. (1975). Southern atlantic coast region. American Birds 29 (1):40-43. (enlace)
- Thompson, W. L., R. H. Yahner, & G. L. Storm (1990). Winter use and habitat characteristics of vulture communal roosts. Journal of Wildlife Management 54:77–83. (enlace)
- Thurber, W. A. (1981). Aerial «play» of Black Vultures. Wilson Bulletin 93:97. (enlace)
- Torres, R. & P. Michelutti (2001). Las aves de ambientes acuáticos del sistema Laguna Mar Chiquita – Bañados del Río Dulce (provincias de Córdoba y Santiago del Estero, Argentina. Bol. Acad. Nac. Cienc. Córdoba 66: 61-73. (enlace)
- Torres, T. F., A. Ríos, E. Schiappacasse, M. Mañez, J. Beruhard & R. Morales (2019). Relevamiento de fauna para la puesta en valor de la Reserva Natural de usos multiples del Cóndor, departamento Pocho, Córdoba, Argentina. Nótulas Faunísticas 265: 1-10. (enlace)
- Travaini, A., A. Rodríguez, O. Ceballos, J. A. Donázar & F. Hidalgo (1995). Roadside raptor surveys in central Argentina. Hornero 14: 64-66. (enlace)
- Trejo, A. & Seijas, S. (2003) . An estimation of road mortality of birds in Nahuel Huapi National Park, northwestern Argentine Patagonia. El Hornero, 018(02), 097-101. (enlace)
- Vallely, A. C. & Dyer, D. (2018). Birds of Central America. Published by Princeton University Press, 41 William Street, Princeton, New Jersey 08540; 570 pp.
- Vera, A. S. (2002). Lista de aves de San Miguel de Tucumán y Yerba Buena. Acta zoológica lilloana, 46(1), 131-135. (enlace)
- Vickery, P. D. (1982). The Autumn migration: August 1–November 30 1981, Northeastern Maritime region. American Birds 36:152–155. (enlace)
- Vides Almonacid, R. (1991). La alteración del boque de Yungas en Tucumán, Argentina y el uso de las aves como indicadores ecológicos para el diseño de zonas de amortiguamiento en áreas protegidas. Tesis Maestría Sc. Programa de Maestría de Manejo de Vida Silvestre para Mesoamérica y el Caribe, Universidad de Costa Rica; 210 pp. (S.I.B.)
- Vides Almonacid, R. (1992). Estudio comparativo de las taxocenosis de aves de los bosques montanos de Sierra de San Javier, Tucumán, Argentina. Bases para su manejo y conservación- Tesis doctoral, Facultad de Ca. Naturales, Universidad Nacional de Tucumán, Argentina; 347 pp.
- Vidoz, F. (1993). Informe de observaciones de avifauna en el Parque Nacional Lago Puelo, en el período julio 1992 y abril 1993. Nota de la DRP Nro. 5 del 17/5/93; 8 pp. (S.I.B.)
- Vidoz, F. (1995). Informe del reconocimiento del valle del río Puelo realizado entre los días 24 y 27 de noviembre de 1994. Nota 49/95 del PN Lago Puelo; 6 pp. y 1 mapa. (S.I.B.)
- Vidoz, F., Berardi, M. & E. Anderson (2011). Informe de Patrulla, Zona Limítrofe Los Hitos Sur – Arroyo Melo, en Parque Nacional Lago Puelo. (S.I.B.)
- von Berlepsch, H. (1908). On the birds of Cayenne, 2. Novitates Zoologicae 15:261–324. (enlace)
- Vuilleumier, F. (1985). Forest birds of Patagonia: ecological geography, speciation, endemism, and faunal history. Ornithological Monographs, 255-304. (enlace)
- Wallace, M. P., P. G. Parker & S. A. Temple (1980). An evaluation of patagial markers for cathartid vultures. Journal of Field Ornithology 51:309–314. (enlace)
- Wallace, M. P. & S. A. Temple (1987). Competitive interactions within and between species in a guild of avian scavengers. Auk 104:290–295. (enlace)
- Ward, P. & A. Zahavi (1973). The importance of certain assemblages of birds as «information centres» for food finding. Ibis 115:517–534. (enlace)
- Wells, J. V. & Wells, A. Ch. (2017). Birds of Aruba, Bonaire, and Curaçao. (1º Ed.). COMSTOCK PUBLISHING ASSOCIATES a division of Cornell University Press Ithaca and London; 474 pp.
- Wetmore, A. (1926). Observations on the birds of Argentina, Paraguay, Uruguay and Chile. United States Nat. Mus. Bull. 133: 1-448. (enlace)
- Wetmore, A. (1956). A checklist of the fossil and prehistoric birds of North America and the West Indies. Smithsonian Miscellaneous Collections 131:1–105. (enlace)
- Wetmore, A. (1962). Systematic notes concerned with the avifauna of Panamá. Smithsonian Miscellaneous Collections 145:1–14. (enlace)
- Wetmore, A. (1965). The Birds of the Republic of Panama. Volume 1. Tinamidae (Tinamous) to Rynchopidae (Skimmers). Smithsonian Miscellaneous Collections 150. Smithsonian Institution, Washington, DC, USA; 483 pp. (157) (enlace)
- Wieczorek, J. (2007). Observation records for Parques. (S.I.B.)
- Wilson, A. S. (1926). Lista de aves del sur de Santa Fe. Hornero 3: 349-362. (enlace)
- Wink, M. (1995). Phylogeny of Old and New World vultures (Aves: Accipitridae and Cathartidae) inferred from nucleotide sequences of the mitochondrial cytochrome b gene. Zeitschrift fuer Naturforschung C 50:868–882. (enlace)
- Wright, A. L., R. H. Yahner & G. L. Storm (1986). Roost-tree characteristics and abundance of wintering vultures at a communal roost in south central Pennsylvania. Raptor Research 20:102–107. (enlace)
- Yahner, R.H., G.L. Storm & A.L. Wright (1986). Winter diets of vultures in southcentral Pennsylvania. Wilson Bulletin 98:157-160. (enlace)
- Yahner, R. H., G. L. Storm & W. L. Thompson (1990). Winter diets of vultures in Pennsylvania and Maryland. Wilson Bulletin 102:320–325. (enlace)
- Yee, D. G., S. F. Bailey & D. Fix (1994). The autumn migration: August 1-November 30 1993. Middle Pacific Coast region. American Birds 48:147–151. (enlace)
- Yepes, J. (1939). Contribución al conocimiento de la Biocenosis del Oeste de la Gobernación del Neuquén. Revista del centro de estudios del doctorado en ciencias Naturales, 2 (5): 65-81. (enlace)
- Yorio P. & Bertelloti, M. (2007). (En A. S. Di Giacomo (editor). Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 340-341. Temas de Naturaleza y Conservación 5. Aves Argentinas/Asociación Ornitológica del Plata. Buenos Aires. (enlace)
- Yzurieta, D. (1995). Manual de reconocimiento y evaluación ecológica de las aves de Córdoba. Ministerio de Agricultura, Ganadería y Recursos Renovables. Córdoba
- Zanotti, M. (2014). Coragyps atratus (Bechstein, 1783). www.ecoregistros.org./61145
- Zapata, A. R. P. (1998a). Contribución al estudio de la avifauna de la región de la Laguna Chasicó, partido de Villarino, provincia de Buenos Aires (Doctoral dissertation, Universidad Nacional de La Plata). (enlace)
- Zapata, A. R. P. (1998b). Aves registradas en la región de la laguna Chasicó, sudoeste de la provincia de Buenos Aires. Natura Neotropicalis 29 (2): 156-161. (enlace)
- Zapata, A. R. P. (2002). Nueva cita sobre la nidificación y crías del Jote cabeza negra (Coragyps atratus) para la provincia de Buenos Aires, Argentina. Nuestras Aves 44: 14-15. (enlace)
-
-
Jote cabeza negra | Conservación
ESTADO DE CONSERVACIÓN SEGÚN BIRDLIFE INTERNACIONAL: PREOCUPACIÓN MENOR
Contaldi y colaboradores, concluyen, que con una población total estimada en 20 millones de individuos, se sabe que hoy sus poblaciones extremas se encuentran en proceso de expansión como consecuencia del impacto de la actividad antrópica en el ambiente a nivel local (mayor oferta de alimento y disminución en la persecución) o global (mayores temperaturas producto del cambio climático) (Contaldi et al., 2019:8)
Esta especie tiene un rango extremadamente grande y por lo tanto no se aproxima a los umbrales de vulnerabilidad bajo criterio de tamaño de la extensión de la población (Extensión < 20.000 km² combinado con una declinación o fluctuación del tamaño de la extensión, extensión/calidad del hábitat, o tamaño de la población y un pequeño número de localizaciones o con fragmentación grave). La tendencia de la población parece estable, por esto esta especie no alcanza los umbrales de Vulnerabilidad bajo el criterio de tendencia poblacional (disminución > 30% en diez años o en tres generaciones). El tamaño de la población es extremadamente grande, por lo tanto esta especie no se aproxima a los umbrales de Vulnerabilidad bajo el criterio de tamaño de la población (<10.000 individuos maduros con una disminución continua estimada > del 10% en diez años o en tres generaciones o con una estructura de población específica). Por estas razones, la especie es evaluada como de “preocupación menor” (Stotz, 1996:142).
En Argentina (S.I.B.):
No Amenazada (NA): según AA y SAyDS, 2008 (Aves Argentinas y Secretaría de Ambiente y Desarrollo Sustentable ). López-Lanús, B., P. Grilli, E. Coconier, A. Di Giacomo y R. Banchs. Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas /AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina. 1 ed. AOP: Buenos Aires, Argentina
No Amenazada (NA): según MADS y AA, 2017 (Ministerio de Ambiente y Desarrollo Sustentable de la Nación y Aves Argentinas ).Categorización de las Aves de la Argentina según su estado de conservación. Informe del Ministerio de Ambiente y Desarrollo Sustentable de la Nación y de Aves Argentinas. 146 p. 2017
No Amenazada (NA): según SAyDS, 2010 (Secretaría de Ambiente y Desarrollo Sustentable ).Categorización de aves autóctonas de Argentina. Lista de aves no registradas, endémicas y exóticas de Argentina (Res 348). 2010
Preocupación Menor (LC): según UICN (API) (Unión Internacional para la Conservación de la Naturaleza ). Lista Roja de la UICN publicada en https://www.iucnredlist.org. Actualización en tiempo real -
Jote cabeza negra | Distribución
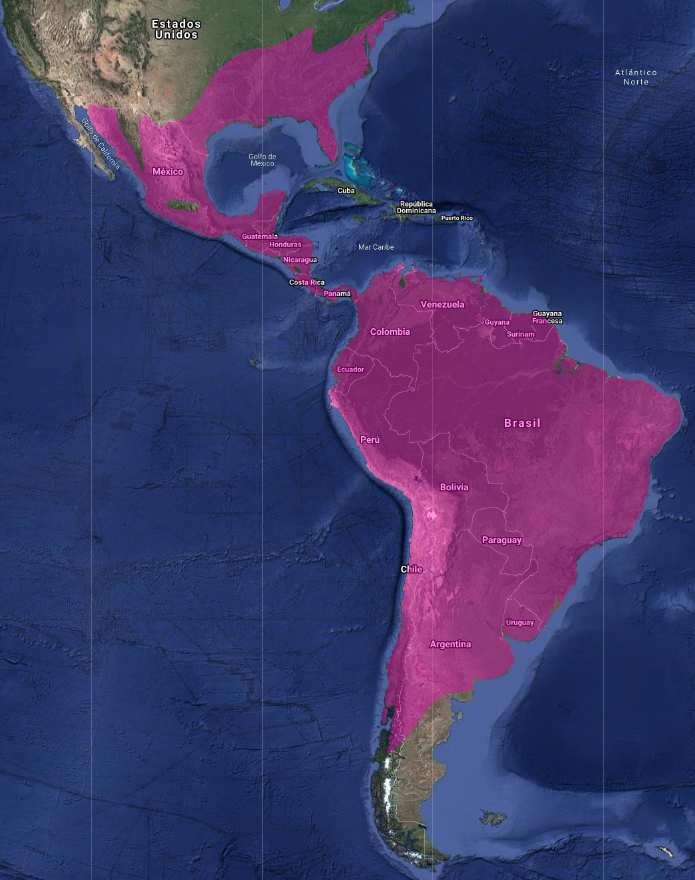
InfoNatura: Animals and Ecosystems of Latin America 2007 Se distribuye por todo el continente americano desde Canadá a Chile y Argentina. Parte oeste de Sudamérica hasta el sur de Chile y Argentina.
En Argentina: Jujuy, Salta, Tucumán, Catamarca, La Rioja, Santiago del Estero, Formosa, Chaco, Misiones, Corrientes, Entre Ríos, Córdoba, Santa Fe, La Pampa, Buenos Aires, San Luis, San Juan, Mendoza, Neuquén, Río Negro, Chubut y Santa Cruz (De la Peña, 1987:44-45); (De la Peña, 1999:37).HÁBITAT
Es la especie más difundida debido en gran parte a su adaptabilidad a diferentes entornos. Habita en lugares de poca vegetación (márgenes de bosques, orillas de ríos, y del mar), bosques, esteros y sabanas. También en zonas urbanas donde se le ve en los basureros.
Ecorregiones: Altos Andes, Bosques Patagónicos, Campos y Malezales, Chaco Húmedo, Chaco Seco, Delta e Islas del Paraná, Espinal, Estepa Patagónica, Esteros del Iberá, Monte de Llanuras y Mesetas, Monte de Sierras y Bolsones, Pampa, Puna, Selva de Yungas, Selva Paranaense. -
Jote cabeza negra | Comportamiento
Muy sociable. Los miembros de la familia mantienen relaciones de larga duración. Los miembros más cercanos de la familia mantienen contacto estrecho a lo largo del año (Rabenold, 1986:32); (Rabenold, 1987b:647); (Parker et al., 1995:395). Viven muchos años, con registros de longevidad de 16 años y 10 meses (Clapp et al., 1982:103)
Durante 3-4 meses después de emplumarse, el nido sirve de lugar de sociabilización diario para jóvenes y adultos (Rabenold, 1986:37). Más tarde, cuando la cría ya es mayor, se encuentran en los posaderos comunales y sitios de alimentación (Rabenold, 1986:37-38).
Los padres alimentan los pichones hasta 6 meses después de emplumarse (Jackson, 1975:603); (Rabenold, 1986:38).
Los juveniles y adultos mantienen contacto estrecho hasta la siguiente estación reproductiva, cuando los adultos se vuelven más agresivos y expulsan a los jóvenes del nido (Stewart, 1983:311).
Una vez separados de sus padres, los juveniles vagabundean entre los posaderos y durante algún tiempo siguen a los adultos (muchas veces intolerantes con ellos), de los cuales aún dependen para alimentarse (Rabenold, 1987a:650-652).
Durante este periodo de vagabundeo, los juveniles aprenden a alimentarse por sí mismos. Demoran mucho tiempo la reproducción (al menos 8 años) (Parker et al., 1995:395).
De manera constante constituyen alianzas, e interactúan entre varias parejas de adultos y sus crías, aparentemente de forma cooperativa en posaderos y carcasas (Parker et al., 1995:395). Existe una fuerte asociación entre el grado de relación genética entre los adultos y cómo ellos interactúan. Individuos que son genéticamente similares es muy probable que utilicen el mismo dormidero la misma noche, así como, el acicalamiento mutuo solo entre parientes (y parejas); se ha observado que las peleas son principalmente entre individuos aparentemente no relacionados (Parker et al., 1995:399).
A menudo, el Jote Cabeza Negra ocupa los posaderos comunales durante varios años, a veces décadas (Prather et al., 1976:668). Algunos son utilizados de forma permanente a lo largo del año, pero otros pueden utilizarse estacionalmente, o de forma temporal, cerca de fuentes de alimento eventuales (Sweeney & Fraser, 1986:24).
Cada individuo utiliza varios posaderos dentro del área de residencia, formando un sistema de posaderos (Stolen, 1996b). A menudo comparte dormideros con el Jote de Cabeza Colorada (Prather et al., 1976:668); (Buckley, 1997:267); (Buckley, 1998:124) y Caranchos (Caracara plancus) (Ng & Jasperson, 1984:214).
Los juveniles que aún son dependientes de sus padres muestran gran fidelidad con los posaderos, pero los mayores prefieren uno solo. Los adultos demuestran fidelidad intermedia (Rabenold 1987b:650).
Son comunes los enfrentamientos en los posaderos comunales. La agresión se dirige a los que no pertenecen al grupo, y los perdedores es poco probable que vuelvan (Rabenold 1987b:651). Los adultos que tienen sus nidos cerca de los posaderos, ganan más enfrentamientos que en posaderos más alejados (Rabenold 1987:651). De esta forma, los adultos que se reproducen localmente pueden controlar, parcialmente, el acceso a los posaderos de individuos que no pertenecen al grupo. Estas agresiones reducen las posibilidades que los adultos sean seguidos a las zonas de alimentación por los extraños y minimizar los costes de compartir alimentos con tales individuos (Rabenold, 1987b:651).
El Jote Cabeza Negra camina y corre bien, a veces con saltitos distintivos y las alas medio abiertas. Según Mahoney (1983, citado en Buckley 2020) corren a velocidades de alrededor de 6.6 km/h. A velocidades >7 km/h cambian de trote a galope, pero solo pueden mantener esta velocidad unos pocos minutos.
Un componente importante en su estrategia de búsqueda de alimento es el vuelo planeado energético-eficiente. Su habilidad para cubrir grandes áreas, de una manera eficiente desde el punto de vista energético, le permite obtener alimentos espacialmente impredecibles, como es la carroña (Parrott, 1970:363).
Generalmente corre 2-3 pasos para tomar el vuelo, aleteando si las termales son suaves. Utiliza el aleteo, normalmente, cuando se desplaza entre los posaderos y los sitios de alimentación conocidos (Kirk & Gosler, 1994:940); (Buckley 1997:267).
Se eleva a gran altura, volando en círculos, aprovechando las corrientes térmicas ascendentes y planea manteniendo las alas en línea horizontal (Pennycuick, 1983:324). Vuelo estable, alternando rápidas batidas de alas con periodos cortos de deslizamiento. En vuelo, cabeza negra, puntas de alas blancas, cola corta en forma de abanico.
Su vuelo es menos eficiente que el de otros buitres ya que las alas no son tan largas por lo que tiene menor superficie para planear. Regurgita cuando se le acercan o es molestado, con lo que disuade a sus predadores y emprende vuelo más fácilmente al disminuir su peso.En tierra tiene el hábito de “urohidrosis”: orina y defeca en sus patas para que se enfríen por evaporación. Tienden a alimentarse durante el día, volviendo a sus dormideros por la noche.
Al carecer de olfato son más gregarios y por lo general dependen de su vista para distinguir animales inmóviles desde cientos de metros de altura. Planea alto aprovechando corrientes térmicas, a menudo con otras aves grandes como Jabirúes (Jabiru mycteria), Tuyuyúes (Mycteria americana) y Chajáes (Chauna torquata) (Di Giácomo & Krapovickas, 2005:227).
Tiene buena relación con el hombre. Puede ser domesticado. Desempeñan importante labor sanitaria por su capacidad de eliminar deshechos de la actividad humana. Se ha demostrado que los microbios del carbunclo y cólera son destruidos en el aparato digestivo del Jote.
Se ha registrado casos de acicalamiento a Caracara plancus (Carancho) en un posadero (Ng & Jasperson, 1984:214) y en carcasas (Haverschmidt, 1977:392); (Silva & Carmo, 2015:22); (Souto et al, 2009:1). El acicalamiento probablemente estrecha las relaciones sociales entre individuos de la especie, pero no se explica cuando se dirige a otras especies. El acicalamiento a Caranchos puede ser un error, o tal vez reduce la agresividad de estos últimos.
El acicalamiento se realiza de forma generalizada (Haverschmidt, 1977:392); (Rabenold, 1986:33); (Stolen, 1996b) entre parejas, familiares, crías y hermanos (Rabenold, 1986:37); (Parker et al., 1995:399), en zonas que no pueden alcanzar con facilidad. Generalmente en los posaderos, pero también en carcasas (Rabenold, 1986:37); (Stolen, 1996b).
Se baña en aguas poco profundas (Rea, 1983: 42); (Sazima, 2011:81)
Suelen ser más tolerantes con otros individuos que nidifican en las cercanías como el Jote Cabeza Colorada (Jackson, 1983); (Stewart, 1974:598); (McHargue, 1977; citado en Buckley, 2020)ALIMENTACIÓNOportunista, se alimenta fundamentalmente de carroña de grandes animales, aunque puede comer todo tipo de cadáveres (Stewart, 1978:79); (Coleman & Fraser, 1987:733); (Rabenold, 1987a:1775); (Buckley, 1996:480). Ocasionalmente consume presas vivas. En un análisis de egagrópilas del área periurbana de Dina Huapi (Río Negro, Argentina), realizado por Ballejo & De Santis, (2013:13), los autores encontraron, además de restos orgánicos, otros sintéticos como restos de bolsa de polietileno, fragmentos de plástico, goma elástica, cartón, fragmento de tela, cuerda, tanza de pesca, fragmento de baldosa, pintura sintética y otros restos indeterminados.
El bulbo olfatorio del Coragyps atratus está menos desarrollado que el del Cathartes aura (Bang, 1964:155); (Bang & Cobb, 1968:58) y a diferencia del Cathartes aura, el Coragyps atratus no utiliza el olfato para localizar los cadáveres (Stager, 1964:55).
Por lo tanto el Cathartes aura es más eficiente al localizar los restos y casi siempre precede a los Coragyps atratus, por lo que dependen de ellos para encontrar alimentos (Stewart, 1978:82); (Houston, 1986:318); (Houston, 1988:405); (Buckley, 1996:473); (Buckley, 1997:267); (Stolen, 1996:43)
Una vez localizado el cadáver, los Coragyps atratus se acumulan rápidamente y desplazan a los Cathartes de la comida (Stewart, 1978:82); (Wallace & Temple, 1987:292); (Houston, 1988:405); (Stolen, 1996:43); (Buckley, 1997:267).
Dado que su estrategia de alimentación se basa en mantener a los otros depredadores a la vista, los C. atratus vuelan más alto que los Catharthes (Stewart, 1978:82); (Houston, 1988:405); (Buckley, 1996:473); (Stolen, 1996:43). Utilizan las térmicas para ganar altitud, y llegan a alcanzar alturas hasta de ≥300 m (Houston, 1988:405).
Los estudios de Rabenold (1987a:1775) y Buckley (1997:267) han demostrado que posado comunal del C. atratus juega un papel importante en su comportamiento de alimentación, dado que puede facilitar la formación de grupos para la búsqueda de alimentos (Rabenold, 1987a:1775); (Buckley, 1996:486), existiendo además, fuertes evidencias que estos grupos funcionan como centros de información (sitio de posado comunal donde se intercambia la información sobre la localización de los alimentos) (Ward & Zahavi, 1973:517).
Los Jotes de Cabeza Negra no pueden abrir carcasas grandes por sí solos, dependen de los grandes carroñeros, que sí lo pueden hacer. Al contrario que el Cathartes aura, el cuál generalmente coloca uno o dos pies sobre la presa para comer, el Coragyps atratus no suele utilizar las patas (Houston, 1988:7).
Cuando la carcasa está suficientemente abierta, pueden extraer las vísceras; esto desemboca en un gran ajetreo de pájaros que rápidamente tiran de ellas para engullir los trozos.
Comen rápidamente (intervalo medio entre picotazos 4.52 s ± 0.8 SE, n = 126 (Houston, 1988:410), mientras que el Cathartes aura es más lento (intervalo medio entre picotazos 6.96 s ± 1.1 SE, n = 131), en parte porque ellos pasan más tiempo en posición alerta. Los Coragyps dependen muy estrechamente de los miembros de su grupo para alertar de la inminente llegada de predadores (Houston, 1988:411), y por ello son capaces de alimentarse más eficientemente.
Sus métodos de alimentación son: 1) picoteando pequeños trozos diseminados alrededor y sobre la carcasa; 2) tirando de tejido muscular laxo o vísceras; 3) Desgarrando tendones de los huesos; 4) desgarrando trozos de piel (Houston, 1988:408).Ocasionalmente atacan y matan animales domésticos, incluso cerdos recién nacidos (Lovell, 1947:131); (Parmalee, 1954:445); corderos (Roads, 1936:219); (Sprunt, 1946:260) y terneros (Parmalee, 1954:445).
Se alimenta sobre el suelo, aunque también a orillas del agua, por carroña o peces pequeños (Jackson et al, 1978:142).
También se los ha citado cazando y comiendo: pichones de Garza Bruja (Nycticorax sp), mofetas y zarigüeyas (McIlhenny, 1939:472), crías de Tortuga Laúd (Dermochelys coriacea) (Mrosovsky, 1971:672). Individualmente atacan pequeños animales, pero los de mayor tamaño lo hacen en grupo. Se los ha citado, comiendo estiércol y material vegetal (McIlhenny, 1945:137). En Surinam, consumen frutos (dañados durante la cosecha) de cocoteros (Cocos nucifera) y Palmera africana (Elaeis guineensis) (Haverschmidt, 1947:210); (Elias & Valencia, 1982:214). En América Central y Sudamérica, generalmente buscan la comida en las ciudades, en vertederos y basura de las calles (Iñigo Elías, 1987:7); (Hill & Neto, 1991:173).
Se lo ha registrado alimentándose de cadáveres de animales silvestres: Mulita grande (Dasypus novemcinctus), Gualacate (Euphractus sexcinctus), Zorro de monte (Cerdocyon thous), Lobito de río (Lontra longicaudis), Coatí (Nasu nasua), Aguará-popé (Procyon cancrivorus), Tapir (Tapirus terrestres), Pecarí de collar (Pecarí tajacu), Ciervo e los pantanos (Blastocerus dichotomus), Guazuncho pardo (Mazama gouazoupira), y Carpincho (Hydrochaeris hydrochaeris). También en cadáveres de grandes aves como Ñandú (Rhea americana), Cigueña americana (Ciconia maguari) y grandes reptiles como Yacaré (Caimán sp.), Curiyú (Eunectes notatus) y Ñacaniná (Hydrodynastes gigas). Peces muertos como Tarariras (Hoplias Malabaricus) y Bagres sapos (Rhamdia quelen). Restos de placentas de pariciones del ganado (Di Giácomo & Krapovickas 2005:227); Huevos de Flamenco austral (Phoenicopterus chilensis) (De la Peña, 2020:14-15). Se han registrado individuos comiendo de bloques de sal (Coleman et al., 1986:281).
No almacenan alimentos pero, visitan de forma repetitiva la misma carcasa durante varios días. Este hábito les permite a aquellos ejemplares que no conocían la localización, descubrirlas con solo seguir a sus con-específicos desde los posaderos (Rabenold, 1987a:1775); (Buckley, 1997:267).VOCALIZACIONESLos Jote Cabeza Negra carecen de siringe y musculatura asociada (Ligon, 1967:19) y por lo tanto solo pueden emitir un número limitado de sonidos.
Los más comunes son silbidos y gruñidos que se escuchan fundamentalmente al comer y volar.
Los “gruñidos” suenan como cuando los perros comen (Baynard, 1914b; citado en Buckley, 2020) o «guff, guff, guff» similar a cuando los perros ladran a la distancia (Saunders, 1906;166).
Los adultos producen un silbido cuando son molestados al comer o en sus nidos (Baynard, 1914; citado en Buckley, 2020).
Sonidos no vocales
Cuando descienden a comer, a menudo lo hacen en ángulo agudo que genera un silbido (Thurber 1981:97).
REPRODUCCIÓNMonógamos; se emparejan para toda la vida (Rabenold, 1986:34); en un estudio realizado en el norte del estado de Carolina, registró que las parejas se mantenían al menos 12 años, y solo se reemplazaban después de la muerte de uno de ellos (Decker et al. 1993:29). Estudios de ADN realizados sobre huellas de 16 familias nucleares, de 32 individuos en reproducción y 36 crías, no revelaron evidencias de paternidad fuera de la pareja; estos resultados indican que las parejas son estrictamente monógamas (Decker et al., 1993:29).
Las parejas mantienen el contacto estrecho a lo largo de todo el año. Suelen posarse cerca del sitio probable antes de la nidificación (Stewart, 1983:310); (Jackson, 1983 y 1988; citado en Buckley, 2020); este comportamiento, el cual puede comenzar unas 4-6 semanas antes de depositar los huevos, puede permitirles valorar la calidad del sitio y la ausencia de elementos perturbadores a su alrededor (Stewart, 1983:310).
En temporada reproductiva, varios autores han descrito vuelos aparentemente de cortejo; un individuo (aparentemente el macho) persigue a la hembra en el aire y periódicamente se lanza hacia ella (Bent, 1937:28-29); (Stolen, 1996b; citado en Buckley, 2020). El macho capta la atención de la hembra volando en círculos, con el cuello estirado hacia delante, mientras exhala profundamente; otra danza ritual consiste en enfrentarse uno a otro en un alarde arriba-abajo con el pico abierto (Rea, 1983; citado en Buckley, 2020); en esta danza, mantienen las alas totalmente desplegadas, alternando con saltos y emisión sonidos similares a “ladridos” (Rabenold, 1986:38)
No construye nido. Deposita directamente los huevos en el lugar elegido y utiliza una gran variedad de sitios de nidificación, como cuevas, huecos oscuros en troncos de árboles y tocones, grietas entre las rocas (Coleman & Fraser, 1989b:21), arbustos y matorrales, edificios abandonados, rascacielos (Hill and Neto 1991:173), etc. (Marchant 1960:360-362); (De la Peña, 2013:88).
En Argentina se reproduce desde fines de julio a mediados de diciembre. Lo hace en huecos de árboles y a veces sobre el suelo en un lugar protegido y oculto. Se encontraron nidos hasta los 6 ó 7 m de altura. Donde depositan los huevos pueden agregar trocitos de corteza o algunos pequeños palitos. En nidos en los pastizales agregaron pastos y trocitos de estiércol seco. Se localizaron nidos en Tucumán, Formosa, Chaco, Misiones, San Juan, Mendoza, La Pampa, Santa Fe, Córdoba, Buenos Aires, Neuquén, Río Negro (De la Peña, 2020:15); (Lucero, 2020:40); (Di Giácomo & Krapovickas 2005:227).
Los sitios de nidificación seguros, frecuentemente son utilizados durante varios años seguidos (Jackson, 1988; citado en Buckley, 2020).
Pone 2-3 huevos, en días consecutivos (Stewart, 1974:596), alternos o con intervalos de 2-3 días (Baynard, 1909; citado en Buckley, 2020). Existen numerosos registros de reemplazo de una nidada perdida (3-4 semanas después) con una nueva puesta (Hill & Neto 1991:175).
La forma de los huevos varía entre ovalado, elíptico o ovoide elongado (raro fusiforme) (Bent, 1937:32). El tamaño de los huevos del C. a. foetens: 2 huevos medidos en Argentina: 74.5 x 45.0 y 72.0 x 47.0, respectivamente (Wetmore, 1962:4).
El color de fondo es variable entre blanco, blanquecino, blanco grisáceo, cremoso, celeste pálido o celeste verdoso pálido, con manchas castañas, violáceas, pardas o negruzcas y pintas grises. Algunos con pocas pintas y manchas. A veces las manchas más concentradas en el polo mayor (Bent, 1937:32-33).
Stewart observó que la incubación comenzaba con la puesta del primer huevo.
En cuanto al periodo de incubación existen numerosas reseñas, pero las más convincentes sugieren que es de 38-39 días (Stewart, 1974:599).
El autor (Stewart, 1974:596) comenta que, los adultos ocasionalmente pierden el rastro de sus huevos durante la noche, con el consiguiente enfriamiento, lo que podría contribuir a la variación en los registros de periodos de incubación. La eclosión puede ser el mismo día, o hasta 3 días después (Stewart, 1974:596).
Ambos padres comparten la incubación por igual, alternándose cada 24 hs (Stewart, 1974:596 y 600).
Mantienen los huevos en la parte superior de los dedos de los pies, dando pequeños pasos y arrastrando los pies debajo de ellos. También utilizan la mandíbula para posicionar los huevos, particularmente para que estén más próximos entre sí; utilizan el pico para recuperarlos cuando salen de esta posición (Stewart, 1974:596).
El primer signo de eclosión es la aparición de una protuberancia cercana al extremo mayor (Bent, 1937:33); la cría extiende la brecha y fuerza la rotura de la cáscara. Este proceso completo puede tardar hasta 24 hs o más.
Nacen cubiertos de abundante plumón de color beige; ojos oscuros y piel desnuda de la cara de color negro (Bent, 1937:32); pesan aproximadamente 70 g (McHargue, 1981:182).
De crecimiento relativamente lento, el peso asintótico se alcanza alrededor de los 60-65 días (McHargue,1981:182); (Coleman & Fraser,1989a: 200-201). Este crecimiento lento puede ser una adaptación a los aportes impredecibles de alimentos debido a las condiciones climáticas (McHargue, 1981:182).
Los autores creen que el mejor predictor de la edad es la medición del ala a los 80 días de edad, ya que es menos variable que el peso (Coleman & Fraser, 1989a:201); (Stewart, 1974:598)
Las alas completamente emplumadas a los 65 días y han desarrollado las plumas del dorso y pecho, pero aún están cubiertos con plumón en zona inferior y cuello. A los 79 días ya están completamente emplumados con algo de plumón presente aún en pecho y vientre (McHargue, 1981:182).
El primer vuelo es alrededor de los 75-80 días (Stewart 1974:598). Una vez que pueden volar, los juveniles suelen permanecer en las cercanías del nido durante alrededor de 10 días antes de abandonar a sus padres.
Los padres cuidan a sus crías continuamente durante los primeros días después de la eclosión (Stewart 1974:596). Mientras aún son pequeños, los cubren con el pecho, pero al crecer son cubiertos con las alas (Stewart, 1974:597).
La frecuencia de estos cuidados disminuye a partir de los 14 días, ya que los polluelos, a esta edad,parecen ser capaces de regular independientemente la temperatura. Stewart observó que en algunos casos el cuidado diurno continuaba hasta los 24 días y hasta 43 días durante las noches, cuando ya los adultos dejan de pasar la noche en el nido.
Cuando son pequeños, son alimentados frecuentemente por regurgitación directamente desde el pico de los padres (15-20 sesiones diarias). En base a sonidos, se cree que también son alimentados por la noche. Alrededor del día 14 comienzan a recibir alimentos sólidos, que aún toman del pico de los adultos. A medida que crecen, los padres pasan largos periodos fuera del nido y alimentan a los pichones solo 2-4 veces al día; solo los días en que las condiciones para el vuelo no son adecuadas (días lluviosos), pueden no alimentarse para nada (Stewart, 1974:597); (McHargue, 1981:182).
Cuando son pequeños, los padres normalmente inician la alimentación extendiendo hacia ellos el pico (Stewart, 1974:597), sin embargo, al crecer los jóvenes solicitan activamente comida a sus padres emitiendo silbidos agudos, moviendo sus cabezas, extendiendo parcialmente las alas y pateando con los pies (McHargue, 1977:328).
A los 21 días ya tienen incorporado el mecanismo de urohidrosis (McHargue, 1981:182).
El periodo de dependencia de los juveniles es largo. La alimentación por los padres continúa hasta 5 semanas después de emplumarse y solicitan comida hasta 15 semanas después (McHargue, 1977:328).
MIGRACIÓN
Residente a lo largo de casi toda su distribución, aunque en invierno muchos individuos se retiran de las zonas extremas del norte y de las grandes altitudes. Por ejemplo, en Missouri, migran hacia el sur desde mediados de octubre a noviembre, y permanecen ausentes hasta febrero o principios de marzo (Robbins & Easterla, 1992:61); (Vickery, 1982:152); (Yee et al., 1994:82).
También realizan movimientos locales, de corta duración, cuando el tiempo es desfavorable y retornan cuando cambian las condiciones climatológicas (Eisenmann, 1963:244); (Skutch, 1969:731); (Wetmore, 1965:161). -
Jote cabeza negra | Taxonomía
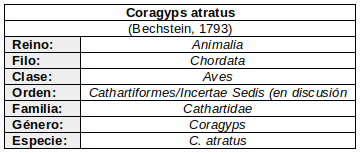
* Los Jotes o Buitres americanos integrantes de esta familia son elevados a un Orden separado: Cathartiformes. Propuesta aceptada por el SACC (South American Classification Committee) Considerado Monotípico por Friedmann (1950:19-30), la American Ornithologyst Union (1957) y Stresemann & Amadon (1979:275). Sin embargo Wetmore (1962:1-4) y Blake (1977; citado en Buckley, 2020: https://doi.org/10.2173/bow.blkvul.01) reconocen 3 subespecies. Su validación requiere futuros estudios de algunas de las características, (no cuantificadas), utilizadas para diferenciar las subespecies (Buckley, 2020: https://doi.org/10.2173/bow.blkvul.01)
SUBESPECIES (Según Wetmore, 1962:1-4)
- C.a. atratus (Bechstein, 1793); “buitre negro de Norteamérica” (México, Tejas y Sur de EEUU)
- C.a. brasiliensis (Bonaparte, 1850); “Buitre negro de América del Sur” (América Central, y norte de América del Sur)
- C. a. foetens (Lichtenstein, 1817); “Buitre negro andino” (Zona andina, desde norte Ecuador, Bolivia, Paraguay, Uruguay, Argentina y tierras bajas de Chile)
-
Jote cabeza negra | Descripción
-
Jote cabeza negra (Coragyps atratus)

Illustrations of the American ornithology of Alexander Wilson and Charles Lucian Bonaparte (1835) NOMBRES COMUNES: Zamuro (Venezuela), Zopilote, Zopilote negro ó Nopo (México y El Salvador), Zopilote o Zote (Honduras), Sucha (Bolivia), Gallote o Gallinazo (Panamá), Zopilote o Zoncho (Costa Rica y Nicaragua), Chulo, Golero o Gallinazo (Colombia), Gallinazo (Perú), Gallinazo Negro (Ecuador), Jote (Chile y Argentina), Cuervo o Buitre negro (Argentina y Uruguay), Yrivu (Paraguay, sur y centro de Brasil), Urubu (Brasil); En guaraní » yryvu»
EN OTROS IDIOMAS
BREVES APUNTES HISTÓRICOS
- A general history of birds. (1821-1828). Latham, John. Winchester [England]: Printed by Jacob and Johnson, for the author: sold in London by G. and W.B. Whittaker. Vol 1: 14-15
- Planches enluminées d’histoire naturelle. (1765-1783). Martinet, François Nicolas, Buffon, Georges Louis Leclerc, comte de, Daubenton, Edme Louis, Daubenton, M. Paris?:s.n., 1765-1783. Vol 2: ilustración 187. (Vautour du Brésil)
- Travels through North & South Carolina, Georgia, east & west Florida, the Cherokee country, the extensive territories of the Muscogulges, or Creek Confederacy, and the country of the Chactaws. Bartram, William. (1793). Dublin,For J. Moore, W. Jones, R. MʾAllister, and J. Rice. Pág. 285. (Vultur atratus)
- Apuntamientos para la historia natural de los páxaros del Paragüay y Rio de la Plata. (1802). Azara, Félix de. Madrid Imp. de la Viuda de Ibarra (1802-1805). Tomo 1: 19 (Iribú)
- Histoire naturelle des oiseaux de l’Amérique Septentrionale : contenant un grand nombre d’espèces décrites ou figurées pour la première fois. (1807). Vieillot, L. P., Bouquet, engraver, Langlois, Pierre Gabriel, printer, Jean Gabriel, , illustrator . A Paris: Chez Desray:1807. Vol 1: 23 (Vultur urubu)
- Dictionnaire d’histoire naturelle. (1816). Paris, Chez Deterville. Vol 12: 401 (Catharista urubu)
- Tableau méthodique des ordres, sous-ordres, familles, tribus, genres, sous-genres et races d’oiseaux. (1831). Lesson, R. P. (René Primevère), Prêtre, J. G., ilustrador. Bruxelles: Chez F.G. Levrault ; Vol 1: 27-28 (Cathartes urubu)
- llustrations of the American ornithology. (1835). Alexander Wilson, Charles Lucian Bonaparte, Turvey, J.; Edimburgh, Frazer & Co.
- Voyage dans l’Amérique méridionale : (le Brésil, la république orientale de l’Uruguay, la République argentine, la Patagonie, la république du Chili, la république de Bolivia, la république du Pérou), exécuté pendant les années 1826, 1827, 1828, 1829, 1830, 1831, 1832, et 1833. (1835-1847). D’Orbigny, Alcide Dessalines d’, Bibron, Gabriel, Blanchard, Emile, Brullé, Gaspard Auguste, Gervais, Paul, Guizot, François, Martius, Karl Friedrich Philipp von, Milne-Edwards, H. (Henri), Montagne, Jean François Camille, Valenciennes, M. Paris: Chez Pitois-Levrault et ce., libraires-éditeurs, rue de la Harpe, no. 81; Vol 4: 31-38 ; Ilustración 1: fig 1 y 2 (Cathartes urubu)
- The birds of America: from drawings made in the United States and their territories. (1840-1844). Audubon, John James, Bowen, John T., lithographer. New York: J.B. Chevalier. Vol 1: 17- 20; ilustración Nº 3 (Cathartes atratus)
- The genera of birds : comprising their generic characters, a notice of the habits of each genus, and an extensive list of species referred to their several genera. (1849). Gray, George Robert, Hullmandel, Charles Joseph, impresor de las ilustraciones. Mitchell, D. W. (David W.), ilustrador. Hullmandel & Walton, impresor de las ilustraciones. London: Longman, Brown, Green, and Longmans. Vol 1: ilustración 1, página 4. (Cathartes foetens)
- Conspectus generum avium. (1850-1857). Bonaparte, Charles Lucian. Lugduni Batavorum :Apud E.J. Brill,. Vol 1: 2. (Vultur atratus)
- American ornithology. (1871). Wilson, Alexander, Baird, Spencer Fullerton, Bonaparte, Charles Lucian. Philadelphia: Porter & Coates. Vol 1-2 (Atlas c-2) (Black vulture)
- Zur Ornithologie Brasiliens: Resultate von Johann Natterers Reisen in den Jahren 1817 bis 1835. (1871). Pelzeln, August von, Natterer, Johann. Wien: A. Pichler’s Witwe & Sohn. Pág. 1 (Cathartes foetens)
- Catalogue of the Birds in the British Museum. (1874-98). British Museum (Natural History). Department of Zoology. [Birds]. London. Vol 1: 24 (Catharistes atratus)
