- Alteff, E. F., G. Gonsioroski, M. Barreiros, L. G. C. O. Torres, A. R. Camilo, H. B. Mozerle, A. E. B. A. Sousa, C. A. B. Medolago. C. Martínez, D. M. Lima, F. K. Ubaid, E. N. Mendonça, B. M. Tomotani & L. F. Silveira (2019). The rarest of the rare: rediscovery and status of the critically endangered Belem Curassow, Crax fasciolata pinima (Pelzeln, 1870). Papéis Avulsos de Zoologia, Museu de Zoologia da Universidade de São Paulo 59:e20195946. https://doi.org/10.11606/1807-0205/2019.59.46 (enlace)
- Argentinas, A. (2017). Categorización de las aves de la Argentina según su estado de conservación. Informe del Ministerio de Medio Ambiente y Desarrollo Sustentable de la Nación y Aves Argentinas. ISO, 690. (enlace)
- Barnett, J. M. & M. Pearman (2001). Lista comentada de las aves argentinas. Lynx Edicions, Barcelona. 164 pp. ISBN 84-87334-32-6
- Bianchini, M. & C. A. Arenas (2014). Primer registro de Muitú (Crax fasciolata grayi) para la Argentina, en el Parque Nacional Chaco, Provincia de Chaco. Bulletin of the Cracid Group Galliformes Specialist Group (GSG IUCN-SSC/WPA). Vol 36:1-4 (enlace)
- BirdLife International (2021). Species factsheet: Bare-faced Curassow Crax fasciolata. Downloaded from https://datazone.birdlife.org/species/factsheet/bare-faced-curassow-crax-fasciolata 22/04/2026
- Bodrati, A. (2005). Las aves del Parque Nacional Chaco. Naturaleza y Conservación, 16: 19. Aves Argentinas. Asociación Ornitológica del Plata. Buenos Aires (enlace)
- Bodrati, A. & M. Lammertink (2011). Una hembra de Mytu (Crax fasciolata) en el Parque Nacional Chaco, Provincia de Chaco, Argentina. Nuestras Aves, 56: 23–25. (enlace)
- Bruno, S. F., Bessa Alves de Carvalho, R. & G. M. Kirwan (2006). Bare-faced Curassow Crax fasciolata in Serra da Canastra National Park, Minas Gerais, Brazil. Cotinga, 26: 80, 86. (enlace)
- Cardiff, S. W. & J. V. Remsen, Jr. (1994). Type specimens of birds in the Museum of Natural Science, Louisiana State University. Occasional Papers of the Museum of Natural Sciences, Louisiana State University, 68: 1–32. (enlace)
- Cebolla Badie, M. (2000). El conocimiento mbya-guaraní de las aves. Nomenclatura y clasificación. Suplemento Antropológico, XXXV(2), 9-188. (enlace)
- Chébez, J. C. (1987). Nuestras aves amenazadas, 21. El Moitú (Crax fasciolata). Nuestras Aves, 14: 17–18. (enlace)
- Chébez, J. C. (1996). Fauna misionera, catálogo sistemático y zoogeográfico de los vertebrados de la provincia de Misiones (Argentina). Monografía No 5. Págs. 320. L. O. L. A. Bs.Aires. 320 pp. ISBN 950-9725-20-X
- Chébez J. C. & H. Casañas (2000). Áreas claves para la conservación de la biodiversidad de la provincia de Misiones (enlace)
- Chébez, J. C. (2008). Los que se van: aves. 1º Ed. Editorial Albatros, Buenos Aires. 416 pp. ISBN: 978-950-24-1255-9
- Collar, N. J., L. P. Gonzaga, N. Krabbe, A. Madroño Nieto, L. G. Naranjo, T. A. Parker & D. C. Wege (1985). Threatened Birds of the Americas. The ICBP/IUCN Red Data Book. Part 2. 3rd edition. ICBP, Cambridge, UK. (enlace)
- Contreras, J. R. (1987). Lista preliminar de la avifauna de la provincia de Formosa, República Argentina. Historia Natural, VII (4): 33-52.
- Contreras, J. R., L. M. Berry, A. O. Contreras, C. C. Bertonatti & E. E. Utges (1990). Atlas ornito-geográfico de la provincia del Chaco. República Argentina. I. No passeriformes. Cuad. Téc. Félix de Azara Nº 1. 164 pág.
- Contreras, J. R., F. Agnolin, Y. E. Davies, I. Godoy, A. Giacchino & E. Ríos (2014). Atlas ornito-geográfico de la provincia de Formosa, República Argentina. 1º Ed. Ciudad Autónoma de Buenos Aires: Fundación de Historia Natural Félix de Azara. 450 p. (enlace)
- de la Peña, M. R. (2006). Lista y distribución de las aves de Santa Fe y Entre Ríos. Monografía (16) L.O.L.A (Literature of Latin America), Buenos Aires.
- de la Peña, M. R. (2013:49). Nidos y Reproducción de las aves argentinas. Serie Naturaleza, Conservación y Sociedad No 8. Ediciones Biológica. Santa Fe. (enlace)
- de la Peña, M. R. (2015). Aves Argentinas. Incluye nidos y huevos. 1º Ed. Santa Fe: Ediciones UNL; CABA: EUDEBA; Vol 1; 496 pp. ISBN 978-987-657-986-5
- de la Peña, M. R. (2020). Aves Argentinas. Descripción, Comportamiento, Reproducción y Distribución (Actualización). Tomo 1: Rheidae, Tinamidae, Anhimidae, Anatidae, Cracidae, Odontophoridae, Phasianidae. Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie).
- del Hoyo, J. & A. Motis (2004). Update chapter. In Curassows and Related Birds. 2nd edition (J. Delacour and D. Amadon), Lynx Edicions, Barcelona, Spain and American Museum of Natural History, New York, NJ, USA. pp. 322–476.
- del Hoyo, J. & N. J. Collar (2014). HBW and BirdLife International Illustrated Checklist of the Birds of the World. Volume 1: Non-passerines. Lynx Edicions, Barcelona, Spain.
- del Hoyo, J. (2020). All the Birds of the world. Lynx Edicions, Barcelona. 965 pp.
- del Hoyo J., Kirwan, G.M. & C. J. Sharpe (2014) Bare-faced Curassow (Crax fasciolata). En: del Hoyo J., Elliott A, Sargatal J, Christie DA y de Juana E (eds) Handbook of the Birds of the World Alive. Lynx Edicions, Barcelona (enlace) http://www.hbw.com/node/53316
- Delacour, J. & D. Amadon (2004). Curassows and Related Birds. Second edition. Lynx Edicions, Barcelona, Spain, and American Museum of Natural History, New York, USA. (enlace)
- Desbiez, A. L. J. & C. Steiner São Bernardo (2011). Density estimates of the Bare-faced Curassow (Crax fasciolata) in the Brazilian Pantanal. Revista Brasileira de Ornitologia, 19 (3): 385–390. (enlace)
- Di Giacomo, A. G. (2005). Aves de la Reserva El Bagual. En Di Giacomo, A. G. y S. F. Krapovickas (Eds.). Historia natural y paisaje de la Reserva El Bagual, provincia de Formosa, Argentina. Inventario de la fauna de vertebrados y de la flora vascular de un área del Chaco Húmedo. Temas de Naturaleza y Conservación 4: 201-465. Aves Argentinas/AOP. Buenos Aires. (enlace)
- Di Giacomo, A. S., M. V. De Francesco & E. G. Coconier (editores). (2007). Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 10-15. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires. (enlace)
- Di Giacomo A. G. (2007:201). Parque Nacional Río Pilcomayo. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 200-201. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
- Di Giacomo A. G. & F. Moschione (2007). Valle Fluvial del Río Paraguay-Paraná. En Di Giacomo, A. S., M. V. De Francesco & E. G. Coconier (editores). (2007). Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 91-92. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
- Di Giacomo A. G. , Gorleri, F. & F. Moschione (2007). Valle Fluvial del Río Paraguay. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 187-188 Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
- Di Giacomo A. G. & E. White (2007). Estancia Guaycolec. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad:193-194. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
- Di Giacomo, A. S. & S. Zalazar (2021). Muitú, el emblema del Chaco Húmedo. Revista Aves Argentinas, 62: 22-25. (enlace)
- eBird (2026): recuperado en abril 2026 de https://ebird.org/species/bafcur1?siteLanguage=es_ES
- EcoRegistros (2026). Muitú (Crax fasciolata) – Ficha de la especie. Accedido de https://www.ecoregistros.org el 21/04/2026.
- Estudillo-López, J. (1977). Observations on the Nocturnal Curassow (Nothocrax urumutum) and other Cracidae. Journal of the World Pheasant Association, 2: 41–49.
- Fernández-Duque, F., Huck, M., Dávalos, V. & E. Fernández-Duque (2013). Estudio preliminar sobre la ecología, el comportamiento y la demografía del Muitú (Crax fasciolata) en la selva en galería del riacho Pilagá, Formosa, Argentina. Hornero, 28 (2): 65-74. (enlace)
- Joseph, L., Slikas, B., Rankin-Baransky, K., Bazartseren, B., Alpers, D. & A. E. Gilbert (1999). DNA evidence concerning the identities of Crax viridirostris Sclater, 1875, and C. estudilloi Allen, 1977. Ornitologia Neotropical, 10 (2): 129–144. (enlace)
- Kirwan, G. M. (2009). Notes on the breeding ecology and seasonality of some Brazilian birds. Revista Brasileira de Ornitologia, 17 (2): 121–136. (enlace)
- Kirwan, G. M., J. del Hoyo, N. Collar, D. A. Christie & C. J. Sharpe (2025). Bare-faced Curassow (Crax fasciolata), version 1.2. In Birds of the World (B. K. Keeney, Editor). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.bafcur1.01.2
- Lepage, D. (2003). Avibase-La base de datos World Bird. Ficha: Pavón Muitú (fasciolata/grayi); recuperado en abril 2026 de https://avibase.bsc-eoc.org/species.jsp?avibaseid=CFC112730C4C332B
- López Lanús, B. (1997). Inventario de las aves del Parque Nacional «Río Pilcomayo» (Formosa, Argentina). Monografía especial Nº 4; 76 pp. Ed. L.O.L.A. (Literature of Latin America)., Buenos Aires, Argentina. ISSN: 0328-1620
- López Lanús, B. & P. Mencia (2017). Diversidad, abundancia y estado de conservación de las aves de los riachos Monte Lindo y Pilagá en el este de la provincia de Formosa, Argentina. Nótulas Faunísticas (segunda serie), 228: 1-29. (enlace)
- Martinez Achenbach, G. (1957). Lista de las aves de la Provincia de Santa Fe. Anal. Mus. Cs. Nat. “ F. Ameghino”, Tomo 1 (1): 1-61 (enlace)
- Moschione, F. (2007). Pilagás III. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 199. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
- Moschione, F. & R. Banchs (2007). Riacho He He. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 201-202. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
- Moschione, F. & H. Casañas (2007). Estancia La Alegría. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 194-195. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
- Nardelli, P. M. (1993). The Preservation of the Alagoas Curassow Mitu mitu, Brazil. Zôo-botânica Mário Nardelli, Nilópolis, Rio de Janeiro, Brazil. 251 pp. ISBN-13: 978-8585579029
- Narosky, T. & D. Yzurieta (2010). Birds of Argentina and Uruguay: a field guide. Vázquez Mazzini Editores, Buenos Aires, Argentina. ISBN 950-99063-6-0
- Navas, J. R. & N. A. Bó (1988). Aves nuevas o poco conocidas de Misiones, Argentina. III. Rev. Mus. Arg. Cienc. Nat. Bernardino Rivadavia, 15 (2):11–37. (enlace)
- Ogilvie-Grant, W. R. (1893). Catalogue of the Game Birds (Pterocletes, Gallinae, Opisthocomi, Hemipodii) of the British Museum. Catalogue of the Birds in the British Museum. Volume 22. Trustees of the British Museum, London, UK. (enlace)
- Pacheco, J. F., G. M. Kirwan, A. Aleixo, B. M. Whitney, A. Whittaker, J. Minns, K. J. Zimmer, P. S. M. Fonseca, M. F. Lima & D. C. Oren (2007). An avifaunal inventory of the CVRD Serra dos Carajás project, Pará, Brazil. Cotinga, 27: 15–30. (enlace)
- Pereira, S. L. & A. J. Baker (2004). Vicariant speciation of curassows (Aves, Cracidae): A hypothesis based on mitochondrial DNA phylogeny. Auk, 121 (3): 682–694. https://doi.org/10.1642/0004-8038(2004)121[0682:VSOCAC]2.0.CO;2 (enlace)
- Pereira, S. L. & A. Wajntal (2001). Estimates of the genetic variability in a natural population of Bare-faced Curassow Crax fasciolata (Aves, Galliformes, Cracidae). Bird Conservation International, 11 (4): 301–308.
- Peters, J. L. (1934). Check-list of Birds of the World. Volume 2. Harvard University Press, Cambridge, MA, USA. https://doi.org/10.5962/bhl.title.14581 (enlace)
- Planqué, R., Vellinga, W. P., Pieterse, S., Jongsma J. & R. de By (Xeno-canto Foundation). Recuperado en abril 2026 de https://xeno-canto.org/species/Crax-fasciolata
- Portes, C. E. B., L. S. Carneiro, F. Schunck, M. de Sousa e Silva, K. J. Zimmer, A. Whittaker, F. Poletto, L. F. Silveira & A. Aleixo (2011). Annotated checklist of birds recorded between 1998 and 2009 at nine areas in the Belém area of endemism, with notes on some range extensions and the conservation status of endangered species. Revista Brasileira de Ornitologia, 19 (2): 167–184. (enlace)
- Ramírez-Llorens, P., E. E. White & M. Rotundo (2003). Sobre algunas aves de la estancia Guaycolec, prov. de Formosa, Argentina. Nuestras Aves, 46: 36-40. (enlace)
- Reichholf, J. H. (1983). Der Bestand an Typenmaterial der ornithologischen Aufsammlungen von J. B. V. Spix in der Zoologischen Staatssammlung München. Spixiana (Supplement 9): 417–423. ISSN 0341-8391 (enlace)
- Rey, N. (2007). Parque Provincial Urugua-í. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 280-281. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/Asociación Ornitológica del Plata, Buenos Aires.
- Robbins, M. B., R. C. Faucett & N. H. Rice (1999). Avifauna of a Paraguayan cerrado locality: Parque Nacional Serranía San Luis, Depto. Concepción. Wilson Bulletin, 111 (2): 216–228. (enlace)
- Saibene, C. A., Castelino, M. A., Rey, N. R., Herrera, J. & J. Calo (1996). Inventario de las Aves del Parque Nacional “Iguazú”, Misiones, Argentina. Monografía, 9. Ed. L.O.L.A. Literature of Latin America, Buenos Aires. 68 pp. ISSN 0328-1620
- Schifter, H., E. Bauernfeind & T. Schifter (2007). Die Typen der Vogelsammlung des Naturhistorischen Museums Wien. Teil I. Nonpasseres. Kataloge der wissenschaftlichen Sammlungen des Naturhistorischen Museums in Wien 20, Aves, Heft 1. Naturhistorischen Museums Wien, Vienna, Austria. (enlace)
- Sclater, P. L. (1875). On the curassows now or lately living in the Society’s Gardens. Transactions of the Zoological Society of London, 9: 273–288. (enlace)
- Servicio de Información de Biodiversidad (SIB). Administración de Parques Nacionales; recuperado en abril 2026 de https://sib.gob.ar/especies/crax-fasciolata
- Sick, H. (1979). Notes on some Brazilian birds. Bulletin of the British Ornithologists’ Club, 99 (4): 115–120. (enlace)
- Sick, H. (1993). Birds in Brazil: A Natural History. Princeton University Press, Princeton, NJ, USA. (enlace)
- Sick, H. (1997). Ornitologia Brasileira. Editora Nova Fronteira, Rio de Janeiro, Brazil. 912 pp. ISBN 85-209-0816-0
- Stotz, D. F., Fitzpatrick, J. W., Parker, T. A., & D. K. Moskovits (1996). Neotropical Birds: Ecology and Conservation. University of Chicago Press, Chicago, IL, USA, and London, UK. 478 pp. ISBN 0-226-77629-8
- Straube, F. C. & M. R. Bornschein (1995). New or noteworthy records of birds from northwestern Paraná and adjacent areas (Brazil). Bulletin of the British Ornithologists’ Club, 115 (4): 219–225. (enlace)
- Tobias, J. A., N. Seddon, C. N. Spottiswoode, J. D. Pilgrim, L. D. C. Fishpool & N. J. Collar (2010). Quantitative criteria for species delimitation. Ibis, 152 (4): 724–746. https://doi.org/10.1111/j.1474-919X.2010.01051.x (enlace)
- Vaurie, C. (1967). Systematic notes on the bird family Cracidae. No. 9. The genus Crax. American Museum Novitates, 2305: 1–20. (enlace)
- Vaurie, C. (1968). Taxonomy of the Cracidae (Aves). Bulletin of the American Museum of Natural History, 138 (4): 131–260. (enlace)
- Warren, R. L. M. (1966). Type-specimens of Birds in the British Museum (Natural History). Volume 1: Non-Passerines. British Museum (Natural History), London, UK. https://doi.org/10.5962/bhl.title.8584 (enlace)
- White, E. (2001). El Muitú (Crax fasciolata) como emblema de una actividad de ecoturismo en el Norte Argentino. Cracid Specialist Group Bulletin, 12: 7–13.
- Zalazar, S. & A. S. Di Giacomo (2019). Consumo de frutos carnosos por el Muitú (Crax fasciolata) en los bosques fluviales del NEA. (enlace)
-
-
Muitú | Conservación
ESTADO DE CONSERVACIÓN EN ARGENTINA: EN PELIGRO DE EXTINCIÓNCrax fasciolata comprende tres subespecies: C. f. pinima (noreste de Brasil), C. f. grayi (este de Bolivia) y C. f. fasciolata (centro y suroeste de Brasil, Paraguay y norte de Argentina) (BirdLife International, 2009). La subespecie C. f. pinima es considerada una entidad taxonómica independiente por BirdLife International y evaluada por separado, encontrándose en estado de Peligro Crítico (Kirwan, 2025).
Las subespecies C. f. fasciolata y C. f. grayi están clasificadas como Vulnerables debido a la pérdida de hábitat y la presión de caza, factores responsables de la extinción local en distintas regiones (Delacour & Amadon, 2004:190). Mientras que C. f. grayi mantiene poblaciones relativamente estables en áreas remotas de Bolivia, la subespecie nominal presenta núcleos aún abundantes en sectores de Brasil, como el Pantanal de Mato Grosso y algunas áreas protegidas, aunque ha desaparecido en São Paulo y es extremadamente rara en Paraná (Kirwan, 2025). En Paraguay, la especie persiste en pocas áreas protegidas y su situación es considerada frágil. (BirdLife International, 2026).
A escala regional, la especie se distribuye desde el este de Bolivia hasta el centro y sur de Brasil, Paraguay y el norte de Argentina. En este último país ha desaparecido de Corrientes, Misiones y Santa Fe, y sus poblaciones actuales son reducidas y fragmentadas (MAyDS & Aves Argentinas, 2017). En Paraguay, aunque previamente se consideraba casi extinta, aún mantiene poblaciones en bosques en galería, donde se ha confirmado su reproducción recientemente.
El tamaño poblacional global no ha sido cuantificado, aunque la especie puede ser localmente común en algunas regiones de Brasil, mientras que en otras áreas es rara o poco frecuente. Estudios de campo muestran densidades variables según el hábitat, con valores relativamente altos en ambientes bien conservados como el Pantanal (Desbiez & São Bernardo, 2011). En contraste, la proximidad a asentamientos humanos reduce significativamente su abundancia (Zalazar et al., 2018), y en Bolivia su estatus ha pasado de relativamente común a raro o poco común (Herzog et al., 2016).
La pérdida de hábitat constituye una de las principales amenazas: entre 2000 y 2019 se perdió aproximadamente el 16% de la cobertura forestal dentro de su rango, con proyecciones de hasta un 30–32% de pérdida en tres generaciones (Global Forest Watch, 2021). A ello se suma la presión de caza, que ha provocado extinción local de la especie y continúa siendo un factor crítico en gran parte de su distribución (Berkunsky & Di Giacomo, 2015).
Como resultado de estos factores, se estima que la población global ha disminuido entre un 20% y un 29% en las últimas tres generaciones, con proyecciones de declive de entre el 30% y el 39% en el futuro cercano (BirdLife International, 2026). Aunque la especie posee un rango de distribución amplio, su tendencia poblacional es decreciente y su conservación depende de la protección de hábitats forestales y del control de la caza.
En contraste, C. f. pinima presenta una situación extremadamente crítica, restringida al Centro de Endemismo de Belém, una de las regiones más deforestadas de la Amazonia, con menos del 25% de su cobertura original remanente. Sus poblaciones están altamente fragmentadas y sometidas a múltiples presiones antrópicas. Tras décadas sin registros, fue redescubierta en 2017 en la Reserva Biológica Gurupi, estimándose una población inferior a 30 individuos (Kirwan, 2025).
A pesar de que Crax fasciolata ha sido considerada previamente como de “Preocupación Menor” debido a su amplia distribución (UICN, 2009), actualmente es reconocida como una especie de alta prioridad de conservación, con tendencias poblacionales negativas vinculadas principalmente a la destrucción del hábitat y la caza (del Hoyo, 1994; BirdLife International, 2009; Stotz et al., 1996:82; Kirwan, 2025).
En Argentina:
En Argentina ha sufrido una marcada retracción, con poblaciones actuales principalmente restringidas a Formosa y, en menor medida, a Chaco (BirdLife International, 2026).
El estado de conservación de Crax fasciolata fasciolata en la provincia de Formosa es considerado crítico a nivel local, debido principalmente a la intensa presión de caza y a los procesos de degradación ambiental, incluyendo la destrucción y contaminación de sus hábitats (Contreras et al., 2014:168). En este contexto, la situación general de la especie es evaluada como de alto riesgo, con un pronóstico desfavorable en ausencia de medidas de control y manejo sostenidas.
No obstante, Formosa ha sido señalada como la provincia que aún alberga las poblaciones más importantes y la mayor disponibilidad de hábitat adecuado para la subsistencia de la especie (Chébez, 1994:409). A pesar de ello, la protección efectiva resulta limitada: fuera del Parque Nacional Río Pilcomayo, sobre cuya población existe escasa información detallada, las áreas protegidas que resguardan a la especie son prácticamente inexistentes. Una excepción relevante es el sector de la Estación de Cría de Animales Silvestres de Guaycolec, adyacente al riacho Pilagá, donde persiste una población significativa (Chébez, 1994:409).
La especie desapareció de Corrientes hacia 1980 debido a la caza y a la transformación del hábitat asociada a la represa de Yacyretá, y actualmente la caza se considera insostenible en algunas regiones. Asimismo, la captura para uso como mascota constituye una amenaza adicional (MAyDS & Aves Argentinas, 2017).
Además de Corrientes, ha desaparecido en las provincias de Misiones y Santa Fe, y sus poblaciones actuales son reducidas y fragmentadas (MAyDS & Aves Argentinas, 2017).Estado de conservación (SIB 2026)
En Peligro (EN): según AA y SAyDS, 2008 (Aves Argentinas y Secretaría de Ambiente y Desarrollo Sustentable). López-Lanús, B., P. Grilli, E. Coconier, A. Di Giacomo y R. Banchs. Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas /AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina. 1 ed. AOP: Buenos Aires, Argentina
En Peligro (EP): según Libro Rojo, 1997 (Fundación para la Conservación de las Especies y el Medio Ambiente). Libro Rojo de Mamíferos y Aves Amenazados de Argentina. García Fernández, J.J.; Ojeda, R.A.; Fraga, R.M.; Díaz, G-B. y R. Baigún. Ed. Dupont. 1997. 221pp. Primera Edición
En Peligro (EN): según MADS y AA, 2017 (Ministerio de Ambiente y Desarrollo Sustentable de la Nación y Aves Argentinas). Categorización de las Aves de la Argentina según su estado de conservación. Informe del Ministerio de Ambiente y Desarrollo Sustentable de la Nación y de Aves Argentinas. 146 p. 2017
En Peligro de Extinción (PE): según SAyDS, 2010 (Secretaría de Ambiente y Desarrollo Sustentable). Categorización de aves autóctonas de Argentina. Lista de aves no registradas, endémicas y exóticas de Argentina (Res 348). 2010
Vulnerable (VU): según UICN (API) (Unión Internacional para la Conservación de la Naturaleza) .Lista Roja de la UICN publicada en https://www.iucnredlist.org. Actualización en tiempo real
En Peligro. Según Categorización de las aves de la Argentina según su estado de conservación. Informe del Ministerio de Medio Ambiente y Desarrollo Sustentable de la Nación y Aves Argentinas. ISO, 690. (2017:16). -
Muitú | Distribución
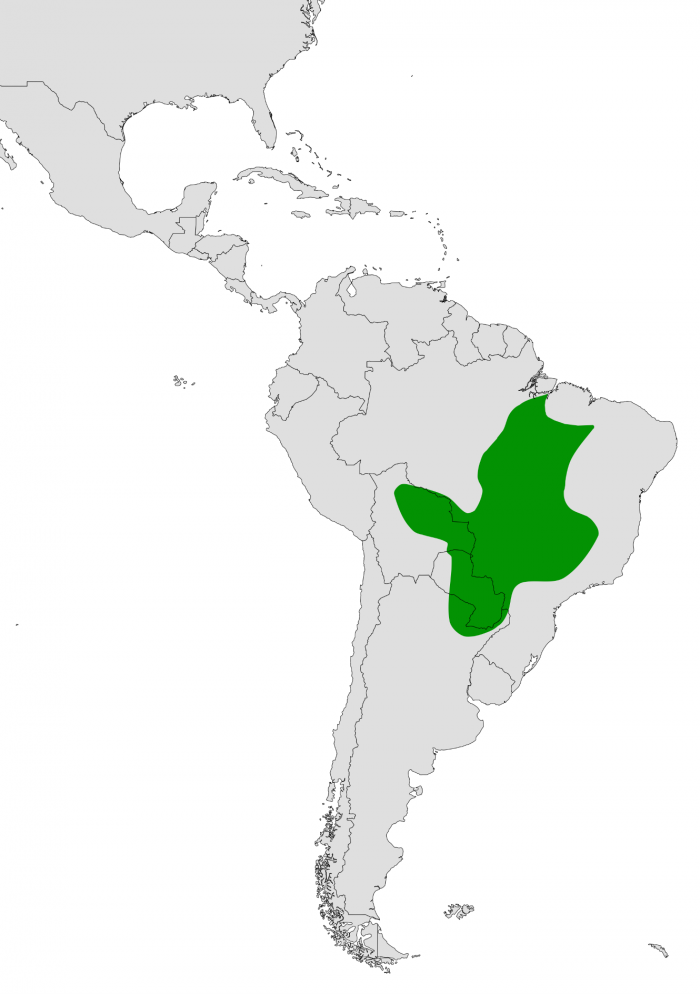
© Planckarte, Mapa de distribución Crax fasciolata. Algunos derechos reservados Según Vaurie (1968:131), Crax fasciolata posee la distribución más amplia dentro de su género. Su rango se extiende desde los estados brasileños de Maranhão y Pará hacia el oeste hasta el río Tapajós, y hacia el sur a través del centro de Brasil, alcanzando Bolivia, Paraguay y el noreste de Argentina (Formosa, Chaco y Misiones). En Bolivia, reemplaza a Crax globulosa al sur de los 15° S, sin superposición aparente. En Brasil, su distribución alcanza aproximadamente el río Tocantins, el río São Francisco en Minas Gerais, y el oeste de São Paulo y Paraná. No obstante, Stotz et al. (1996:303) la citan únicamente para Paraguay y Brasil.
Información más reciente amplía y confirma este patrón de distribución, indicando que la especie se encuentra en Brasil al sur del Amazonas, desde el río Tapajós hacia el este hasta Pará y Maranhão, y hacia el sur a través de Mato Grosso, Bolivia amazónica, Paraguay y el Chaco del norte-centro de Argentina, incluyendo el norte de Misiones. En el sector oriental, también ha sido registrada en el oeste de Minas Gerais, el oeste de São Paulo (al menos históricamente) y el oeste de Paraná (Winkle et al., 2025). Se han documentado extinciones a nivel local a lo largo de su rango, tanto en la subespecie nominal como en C. f. grayi (Delacour & Amadon, 2004:190). Asimismo, resulta incierto si la especie, en particular C. f. fasciolata o C. f. pinima, ocupó previamente regiones del nordeste de Brasil, como Alagoas; sin embargo, parece estar extinta en la provincia de Santa Fe, Argentina.
En cuanto a la subespecie nominal, C. f. fasciolata, esta se distribuye desde el Brasil central al sur de la Amazonia hasta São Paulo y Mato Grosso, extendiéndose por Paraguay y el norte de Argentina. En este último país ha sido registrada en Misiones, el noreste de Corrientes, Santa Fe (donde posiblemente esté extinta), y el este de Chaco y Formosa (Chébez, 2008:230; Bodrati & Lammertink, 2011:23; Stotz et al., 1996:151).
SUBESPECIES
Se reconocen tres subespecies:
* Crax fasciolata pinima (extremo noreste), posiblemente extinta, es extremadamente rara y se conoce a partir de solo siete ejemplares recolectados entre 1830 y 1907.
* Crax fasciolata grayi (Bolivia),
* Crax fasciolata fasciolata (nominal) (resto del rango).La variación geográfica de Crax fasciolata afecta principalmente a la hembra y es altamente marcada (Vaurie, 1968).
Esta variación presenta un patrón aparentemente clinal, aunque las conexiones entre poblaciones no están completamente esclarecidas.
Las diferencias entre subespecies incluyen variaciones en el ancho del barrado, la coloración de la cresta, la tonalidad dorsal y los patrones de las partes inferiores. En términos generales, pinima presenta una coloración más oscura, grayi es más pálida, y la forma nominal muestra características intermedias, con una tendencia hacia grayi en el sur de su distribución.HÁBITAT
La mayoría de los registros de Crax fasciolata provienen de hábitats forestales cerrados (72%, n = 137), mientras que una proporción menor corresponde a praderas abiertas (28%, n = 53). En estos últimos ambientes, los individuos suelen encontrarse próximos a bordes de bosque, y en llanuras aluviales se restringen a pequeños fragmentos forestales. Estos patrones indican que los hábitats boscosos son fundamentales para la especie (Desbiez & Steiner, 2011:387; Contreras et al., 2014:169).
La especie habita selvas húmedas siempreverdes (Stotz et al. 1996:150), bosques semicaducifolios y selvas en galería, en ocasiones con presencia de bambú, alcanzando al menos los 700 msnm de altitud en Bolivia y los 800 msnm en el este de la Amazonia brasileña. Frecuentemente se la registra en bordes de bosque y en proximidad a cursos de agua, y ocasionalmente deambula por playas fluviales, principalmente durante las primeras horas de la mañana o al atardecer.
Se trata de un ave típicamente forestal, de hábitos mayormente arbóreos, aunque desciende al suelo para alimentarse. Según Chébez (1994:409), “parece preferir como hábitat las selvas umbrías que bordean los cursos de agua o cubren las islas, tanto de riachos como de grandes ríos”. Asimismo, ha sido registrada en diversos ambientes, incluyendo el Bosque Atlántico, el Chaco, la sabana mesopotámica y la selva Paranaense (Stotz et al., 1996:150; Barnett & Pearman, 2001:35).
Es una especie no migratoria. Se ha documentado su carácter residente en el Pantanal de Mato Grosso, Brasil, y probablemente esta condición se extiende a lo largo de toda su área de distribución.ARGENTINA: Distribución geográfica por provincias
En Argentina, Crax fasciolata presenta una distribución histórica amplia pero actualmente fragmentada y en retroceso. La especie ha desaparecido o se encuentra probablemente extinta en varias provincias, persistiendo con mayor seguridad en Formosa y, de manera más limitada, en sectores del Chaco. Su conservación depende estrechamente de la disponibilidad de hábitats forestales bien conservados y de la reducción de las presiones antrópicas, especialmente la caza.
Está representada por dos subespecies:- Crax fasciolata fasciolata (von Spix, 1825), registrada en Formosa, Chaco y Misiones
Históricamente, C. f. fasciolata se distribuía en Argentina en las provincias de Formosa, Chaco, Santa Fe, Corrientes y Misiones (Martínez Achenbach, 1957:18; Olrog, 1979:78; Partridge en Chébez, 2008:233; Bodrati & Lammertink, 2011:24; Chébez, 2008:231). Sin embargo, en Santa Fe, Corrientes y Misiones no existen registros adecuadamente documentados en las últimas décadas, por lo que su presencia actual es incierta o podría considerarse localmente extinta (de la Peña, 2006:125; Chébez, 2008:231). En contraste, las poblaciones más importantes del país se concentran en la provincia de Formosa (White, 2001; Ramírez Llorens et al., 2003:38; Chébez, 2008:231; Bodrati & Lammertink, 2011:24).
Dentro del sistema federal de áreas protegidas, la especie cuenta con poblaciones confirmadas únicamente en el Parque Nacional Río Pilcomayo (Formosa). Su inclusión inicial en esta área se basó en el hallazgo de plumas pertenecientes a una hembra en 1993 (Heinonen Fortabat et al., 1995:999; López-Lanús, 1997:32), aunque posteriormente fue redescubierta en las selvas de ribera del río Pilcomayo (E. White, com. pers.). En el Parque Nacional Chaco, en cambio, fue considerada extinta durante varias décadas debido a su ausencia y a testimonios de pobladores locales (Bodrati, 2005:15). Testimonios históricos indican que la especie era frecuente en el monte fuerte y en las selvas del río Negro hasta la década de 1960, pero la intensa presión de caza provocó una drástica disminución poblacional hacia fines de esa década, seguida de una prolongada ausencia de registros, estimada en aproximadamente 30 años (Bodrati & Lammertink, 2011:24).- Crax fasciolata grayi (Ogilvie-Grant, 1893), presente en la provincia de Chaco (de la Peña, 2020:289).
Situación por provincias
Chaco (Citas)
La situación de la especie en la provincia de Chaco es compleja y altamente comprometida, con escasas áreas de presencia actual (Contreras et al., 1990; Chébez, 2008:231). No obstante, existen registros recientes en el Parque Nacional Chaco, incluyendo la observación y documentación fotográfica de una hembra el 5 de agosto de 2009 en el cauce seco del río Negro (26°48’S, 59°36’O) (Bodrati & Lammertink, 2011:24), así como el avistamiento de otro ejemplar en agosto de 2012 en inmediaciones del Centro de Interpretación del Parque Nacional Chaco (Bianchini & Arenas, 2014:2). También se han reportado registros en el este de la provincia, incluyendo la selva del río de Oro y el río Guaycurú, con observaciones de grupos de hasta ocho individuos, parejas y familias con crías (Braslavsky en Chébez, 2008:231). Datos adicionales indican su presencia en el noreste provincial (Contreras et al., 1990) y registros confirmados hasta los 61°30’ O (Contreras et al., 2014:168). Asimismo, se han documentado evidencias reproductivas (huevos y nidos) en el Parque Nacional Chaco (Di Giácomo & Zalazar, 2021:25). Sin embargo, algunos registros recientes aún carecen de validación (SIB, 2026).Corrientes (Citas)
En Corrientes, la especie cuenta con registros puntuales en la isla de Apipé y en Villa Olivari (Chébez, 2008:231), basados principalmente en comunicaciones personales, sin confirmaciones recientes, donde habría persistido hasta la década de 1960 en áreas posteriormente transformadas por la construcción de grandes represas.Formosa (Citas)
La provincia de Formosa constituye el principal núcleo poblacional de la especie en Argentina. Se la ha registrado en el este provincial, particularmente en ambientes asociados al río Paraguay y sus afluentes, como los riachos Pilagá y He Hé, Campo Bouvier y el río Pilcomayo (Chébez, 1994; Chébez, 2008:231). En áreas como la estancia Guaycolec y el ex Refugio Privado Campo Bouvier persisten poblaciones residentes y reproductivas.
Diversos estudios indican que la especie es cada vez más rara en la provincia, con registros concentrados en la mitad oriental y especialmente en el valle fluvial del río Paraguay, donde las condiciones de accesibilidad limitada y la presencia de bosques densos favorecen su persistencia, aunque en forma relictual (Contreras et al., 2014:168). Se han documentado registros en múltiples áreas clave para la conservación (AICA’s), incluyendo los departamentos Pilcomayo, Formosa y Laishi (Di Giácomo et al., 2007:182/187), así como poblaciones nidificantes en la estancia Guaycolec (Di Giácomo & White, 2007:193; Fernández-Duque, 2013:70).
Existen además numerosos registros de campo, incluyendo 43 avistajes entre 1997 y 2001 en la selva en galería del riacho Pilagá, con observaciones de hembras con pichones (Ramírez-Llorens et al., 2003:38). También se han registrado áreas de alta densidad en sectores bien conservados de los riachos Pilagá y Monte Lindo (López-Lanús & Mencia, 2017:5). Sin embargo, en algunas áreas como la Reserva El Bagual, la especie es considerada probablemente extinta (Di Giácomo et al., 2005:433).Misiones (Citas)
La presencia de Crax fasciolata en Misiones ha sido históricamente controvertida y basada en escasos registros, muchos de ellos dudosos. El único ejemplar que confirma su presencia corresponde a una hembra colectada el 11 de mayo de 1959 en el arroyo Urugua-í (Navas y Bó, 1988; Chébez, 2008:231). Otros registros incluyen observaciones en Cerro Corá (De Basaldúa, 1901), Alto Paraná (Bertoni, 1913) y el arroyo Uruzú en 1986, aunque este último fue posteriormente puesto en duda (Chébez & Casañas, 2000).
Adicionalmente, existen menciones para localidades como Montecarlo, Río Paranay y Colonia Delicia, aunque también de carácter incierto (Chébez & Casañas, 2000). La especie figuró en una lista anónima del Parque Nacional Iguazú en 1983, pero fue excluida en ediciones posteriores (Saibene et al., 1996:29). En conjunto, la evidencia sugiere que la especie estaría actualmente extinta en Misiones, donde su distribución habría sido marginal y restringida a selvas ribereñas (Chébez & Casañas, 2000; Chébez, 2008:231; Rey, 2007:280).Santa Fe (Citas)
En la provincia de Santa Fe existe un único registro visual en el extremo nordeste (Dpto General Obligado, Campo Urdániz), sin fecha precisa (Martínez Achenbach, 1957; en Chébez, 1987:19). La especie se considera actualmente extinta o próxima a la extinción en esta provincia (Chébez, 2008:231). -
Muitú | Comportamiento
Comportamiento generalmente cauteloso y evasivo frente a la presencia humana. Se ha observado que los individuos reaccionan rápidamente ante posibles disturbios, alejándose con prontitud mediante el ascenso a los árboles desde el suelo o desplazándose a través del dosel, mientras emiten un silbido de alarma fuerte y explosivo (Bodrati & Lammertink, 2011:25).
En condiciones habituales, la especie se observa en parejas. Cuando es sorprendida, especialmente en presencia de crías, puede optar por huir corriendo por el suelo; alternativamente, se desplaza entre la vegetación emitiendo un silbido suave y agudo, ocultándose entre el ramaje hasta desaparecer de la vista (López-Lanús, 1997:32). Estas conductas evidencian una estrategia antipredatoria basada en la evasión rápida y el uso del denso entorno forestal como refugio.VOCALIZACIONESEl repertorio de Crax fasciolata se caracteriza por una notable diversidad de emisiones sonoras, que cumplen funciones comunicativas en distintos contextos comportamentales (Kirwan et al., 2025).
Canto: Consiste en una serie de profundos sonidos guturales descritos como “hm-hm-hm hm-hm hm”, que ocasionalmente pueden ser emitidos desde el suelo (Delacour & Amadon, 2004:192). En situaciones de irritación, se ha descrito una vocalización adicional consistente en un silbido agudo, “PSEEEEew” (Sick, 1993:198).
Llamadas: Las llamadas de alarma incluyen: silbido fino similar a “psiiü”, así como “ladridos” de tipo “kwoa”, “wãâu” o “u-ú” o variantes como “waayoo”, a menudo seguidas de un sonido susurrado, en contextos de alerta. especialmente cuando los individuos son perturbados durante la noche en sus perchas (Sick, 1993:191; 1997:280). En estas situaciones de sobresalto nocturno en el dormidero, se reporta también un gruñido fuerte emitido tras la vocalización “wãu”, junto con un sonido susurrado posterior a “waayoo” (Sick, 1993:198).
Asimismo, se ha descrito un silbido prolongado de tono medio, “SFEEeouuw”, que puede transformarse en una emisión más breve, enérgica y aguda, tipo estornudo (“kseuw”), a veces repetido y con inflexión descendente (del Hoyo & Motis, 2004).
El canto puede escucharse durante la noche y se intensifica en las primeras horas tras el amanecer, prolongándose generalmente durante la primera hora del día, aunque en ocasiones excepcionales puede registrarse hasta aproximadamente las 10:00 h (Robbins et al, 1999:219).ALIMENTACIÓNDieta omnívora con predominio de componentes vegetales. Según Kirwan et al. (2025), consume principalmente frutos, generalmente aquellos caídos de los árboles y recolectados en el suelo. Complementa su dieta con semillas, flores e invertebrados (Desbiez & Steiner, 2011:389), así como con brotes, hojas, crustáceos, insectos, miriápodos y moluscos (de la Peña, 2020:287).
Se ha registrado el consumo de hojas de «Redonditas de agua» (Hydrocotyle s.p.) (Bodrati & Lammertink, 2011:25) y la visita a salinas para la ingesta de suelos ricos en sales minerales. Asimismo, se ha documentado su alimentación en flores de especies arbóreas del género Tabebuia (Lapacho), en asociación con otras cracidae como la Charata (Ortalis canicollis), Pava goliazul (Pipile cumanensis) y la Pava ventrirrufa (Penelope ochrogaster) (Collar et al., 1985:125).
En ambientes forestales del norte argentino, se han identificado diversos recursos frutales en su dieta, incluyendo «Chrysophyllum gonocarpum« (Aguaí – aguay – awaí), «Plinia trunciflora« (Guapurú – Ibá porú – îwa purú – Yvapuru), «Syagrus romanzoffiana« (Pindó – ybá pytá), «Eugenia uniflora» (Iba porei – Iba poroití – Nangapirí – Ñangapirî – Pitanga), «Eugenia repanda» (Arrayán), «Inga uruguensis« (Ingá) y «Ocotea diospyrifolia« (Laurel ayuí – Laurel kirimít – Laurel negro) (Zalazar & Di Giacomo, 2019; Zalazar et al., 2019).
Desde el punto de vista ecológico, se ha observado que las semillas ingeridas suelen resultar inviables tras expulsarlas; sin embargo, las de «Syagrus romanzoffiana» pueden atravesar el tracto digestivo intactas, lo que sugiere un potencial papel en la dispersión de esta especie vegetal.
Se alimenta sola o en parejas, principalmente en el suelo o en sus proximidades.
REPRODUCCIÓNSe han registrado pichones en diciembre en Paraguay, mientras que en Formosa (Argentina) se documentó un nido con huevos a finales de noviembre (Kirwan et al., 2025). En Brasil, se han observado adultos acompañados de crías en distintos periodos del año, incluyendo julio y agosto en Minas Gerais (Bruno et al., 2006:86), noviembre en el sur de Mato Grosso y finales de diciembre en Goiás (Kirwan, 2009:122).
El sistema de apareamiento se considera, de manera general, monogámico; sin embargo, observaciones realizadas en el Pantanal de Mato Grosso (Brasil) sugieren la posible existencia de comportamientos de tipo lek en los machos (Desbiez et al., 2011:387).
Los nidos se ubican en árboles, habitualmente a alturas de aproximadamente 3,5 a 4 m sobre el nivel del suelo, y suelen encontrarse bien ocultos entre enredaderas o vegetación trepadora. La estructura consiste en una plataforma sólida construida con palitos, ramitas, tallos herbáceos y hojas, con un revestimiento interno de hojas frescas. Se han descrito plataformas de aproximadamente 40 cm de ancho y 10 cm de profundidad (de la Peña, 2020:287).
En Formosa se localizaron nidos y/o pichones durante los meses de noviembre, diciembre y enero (de la Peña, 2020:287). En Pirané (Dto. Pirané, Formosa) se registraron dos nidos: uno el 22-11-1972 con dos huevos y otro el 02-12-1973 con un huevo (localizado por Juan Ortega). Ambos se encontraban en árboles a 4 m y 3,50 m del suelo, respectivamente, ocultos por enredaderas. Las plataformas estaban constituidas por palitos y hojas, con dimensiones de 40 × 60 cm en uno de los casos y aproximadamente 60 cm de diámetro en el otro, con alturas de 20 cm y profundidades de 10 a 20 cm.
Los huevos son de forma elíptica, de color blanco y con cáscara rugosa. Se ha descrito que la puesta habitual es de dos huevos.
El período de incubación se estima en aproximadamente 28 a 32 días en condiciones de cautiverio, siendo realizado exclusivamente por la hembra (Delacour & Amadon, 2004:190).Al momento de la eclosión, los pichones presentan un peso aproximado de 90 g. Su desarrollo temprano es relativamente rápido, alcanzando cerca de 400 g a los 28 días y aproximadamente 1.260 g a los 93 días de vida (Delacour & Amadon, 2004).
Las crías pueden permanecer junto a los adultos durante varios meses tras la eclosión. En Formosa, se han observado hembras acompañadas de pichones en distintas ocasiones: en Estancia Guaycolec, dpto. Formosa, se registró el 06-01-1999 una hembra con dos pichones y el 05-01-2000 otra hembra con un pichón (Ramírez Llorens et al., 2003:37).
-
Muitú | Descripción

© Bernard Dupont. Macho. Pouso Alegre, Transpantaneira, Poconé, Mato Grosso, Brasil. Julio 2019. Algunos derechos reservados 
© Bernard Dupont. Hembra. Transpantaneira, Poconé, Mato Grosso, Brasil. Junio 2019. Algunos derechos reservados (Kirwan et al., 2025; Vaurie, 1968:238)
Esta especie constituye el único representante del género «Crax» que presenta piel desnuda de color negro alrededor del ojo, rasgo diagnóstico de gran valor taxonómico (Kirwan et al., 2025). Asimismo, se distingue de las demás especies del género por presentar una menor cobertura de plumas en las lores y en la región facial, característica que acentúa la visibilidad de la piel desnuda (Vaurie, 1968:238).MACHO
El macho se caracteriza por un plumaje predominantemente negro, incluyendo la cresta rizada, con excepción de la región ventral blanca y de las timoneras externas, que presentan un estrecho borde apical blanco. La cera es de color amarillo, con un desarrollo incipiente del tubérculo y ausencia de carúnculas; no obstante, esta estructura resulta más robusta que en el «Crax alector«.
Presenta un plumaje negro uniforme con un leve brillo verdoso-azulado, particularmente en el manto. El vientre y las plumas tibiales son blancos, generando un contraste marcado con el resto del cuerpo. Ocasionalmente pueden observarse algunas plumas blancas en los muslos, posiblemente más frecuentes en individuos jóvenes, así como pequeñas puntas blancas en la mayoría de las timoneras; la cola, en conjunto, presenta una terminación blanca característica (Vaurie, 1968:238).
En el macho juvenil, el plumaje es similar al del adulto, aunque presenta tonalidades amarillentas en la piel desnuda situada por debajo del ojo.HEMBRA
Las hembras presentan una marcada variabilidad en el patrón de barrado del dorso, la cola, el pecho y la región superior del vientre, así como en la tonalidad general de las partes inferiores. En este sentido, Vaurie (1968:238) destaca que el patrón de barrado dorsal y ventral muestra una considerable variación geográfica, lo que sugiere una diferenciación poblacional dentro de la especie.
La cresta está compuesta principalmente por plumas blancas con ápices negros rizados y, en ocasiones, con bases oscuras; en conjunto, se encuentra moderadamente desarrollada, aunque en muchas hembras presenta una proporción de blanco mayor que en otras especies del género (Vaurie, 1968:238). El cuello, la parte superior del pecho, el manto, el dorso, las alas y la cola presentan coloración negra o negro grisácea, con un barrado blanco generalizado, excepto en el cuello, que es uniforme. La cola exhibe aproximadamente doce barras blancas estrechas, y todas las timoneras, excepto el par central, presentan puntas blancas de aproximadamente 2,5 cm. Las partes inferiores, salvo el pecho superior, son de color ocráceo intenso a leonado, mientras que el pecho muestra un patrón barrado oscuro sobre fondo ocráceo. Se ha sugerido que esta tonalidad es más intensa en individuos jóvenes.El pico es oscuro, con la punta de color córneo; en el macho, la cera es de color amarillo brillante y ligeramente engrosada, mientras que en la hembra tanto el pico como la cera presentan tonalidades oscuras. El iris es marrón oscuro en los machos y más claro en las hembras. La piel facial desnuda que rodea el ojo es negra en ambos sexos.
Las patas muestran dimorfismo sexual: los tarsos y dedos son de color marrón grisáceo oscuro en los machos, mientras que en las hembras presentan una tonalidad rosada pálida (Kirwan et al., 2025).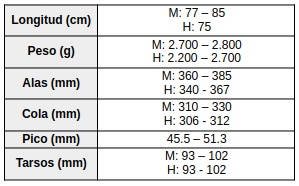
(Vaurie, 1968:238); (de la Peña, 2020:287); (Kirwan et al., 2025); (Delacour & Amadon, 2004:190) Especies similares
Dentro de su amplia área de distribución, esta especie resulta inconfundible. En particular, no coexisten otras especies de crácidos predominantemente negras con pico mayoritariamente amarillo. Aunque los machos son morfológicamente similares a ambos sexos del «Crax alector«, no existe solapamiento geográfico entre ambas especies, lo que facilita su identificación en campo (Kirwan et al., 2025). -
Muitú | Taxonomía
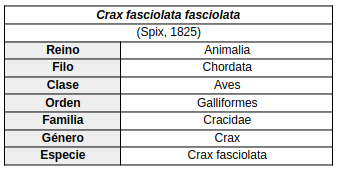
Se reconocen tres subespecies de Crax fasciolata: (Kirwan et al., 2025)
- Crax fasciolata fasciolata (Spix, 1825) (Especie nominal)
Subespecie politípica (del Hoyo, 2020:45). Incluye como sinónimos Crax sclateri (Gray, 1867) y Crax sulcirostris (Ihering, 1898).
Se distribuye en el centro y suroeste de Brasil (al sur del Amazonas), Paraguay y el norte de Argentina (Formosa, Chaco, Corrientes y Misiones) (BirdLife International 2009).
La nomenclatura Crax sclateri (pava de Sclater), descrita a partir de dos ejemplares erróneamente atribuidos a México, es considerada sinónimo de la subespecie nominal (Pinto, 1935; Vaurie, 1967:8). - Crax fasciolata pinima (von Pelzeln, 1870)
Subespecie monotípica. El holotipo corresponde a una hembra adulta recolectada por Johann Natterer el 24 de febrero de 1835, conservada en el Museo de Historia Natural de Viena (NMW 37.949). Este ejemplar fue descrito a partir de un manuscrito inédito de Natterer (Shifter et al., 2007:86). La validez taxonómica de la forma fue inicialmente cuestionada por Sclater (1875:281), siendo posteriormente defendida por Hellmayr (1906:681). Pelzeln y Lorenz (1888:50) la reconocieron como material tipo procedente de Brasil.
Crax incommoda (Sclater, 1872) es considerado sinónimo de C. f. pinima (Peters, 1934:11). C. f. pinima fue elevada a rango específico por del Hoyo y Collar (del Hoyo & Collar, 2014), con base en el ejemplar AMNH 6482, una hembra significativamente más pequeña que las de fasciolata, con diferencias morfométricas según Tobias et al., (2010).
Se caracteriza por una coloración dorsal más oscura, barrado muy reducido (casi ausente en la cola) y partes inferiores notablemente más pálidas (Vaurie, 1967:7).
Su distribución se restringe al extremo noreste de Brasil, específicamente al noreste de Pará (al este del bajo río Tocantins) y al norte de Maranhão, sin registros confirmados en estado silvestre entre finales de la década de 1970 y 2013 (Portes et al., 2011:171; Alteff et al., 2019:1-6). Se ha señalado su extrema rareza, con individuos aún presentes en libertad, pero en densidades muy bajas.
En cuanto a la identificación, el macho es predominantemente negro, con cresta de plumas alargadas rizadas y cloaca blanca, piel facial desnuda negra y cera amarilla en la base del pico, diferenciándose del macho de C. f. fasciolata por su menor tamaño. La hembra presenta dorso oscuro con barrado pálido estrecho, cresta bicolor (blanca y negra) y partes inferiores beige pálido a blanquecino, siendo notablemente más pequeña y más oscura que la hembra de la subespecie nominal (Vaurie, 1968:238; Delacour & Amadon:190/191). - Crax fasciolata grayi (Ogilvie-Grant, 1893)
Presenta la misma distribución general que la subespecie nominal en el centro y suroeste de Brasil, Paraguay y el norte de Argentina (Formosa, Chaco, Corrientes y Misiones) y este de Bolivia.
Algunos machos pueden presentar abdomen beige muy pálido (Sick, 1979:117). En las hembras, la cantidad de blanco en la cresta muestra alta variabilidad individual, mientras que, en comparación con la subespecie nominal, presentan un plumaje general más pálido, barrado más amplio y menos denso, y una mayor extensión de manchas ocres en las cobertoras alares (Vaurie, 1968:239; Delacour y Amadon, 2004:191).
La forma Crax estudilloi ha sido interpretada inicialmente como un posible híbrido; sin embargo, actualmente se considera poco probable que involucre a C. fasciolata, habiéndose propuesto como posibles taxones parentales a Crax daubentoni y Crax alberti (Joseph et al., 1977:138; Allen et al., 1977; Estudillo-López, 1977). El holotipo corresponde a un esqueleto completo con plumas de un macho adulto, depositado en el Museo de Ciencias Naturales de la Universidad Estatal de Luisiana (LSUMZ 140000) (Cardiff & Remsen Jr., 1994:2).
Adicionalmente, la subespecie C. xavieri, descrita a partir de un macho en cautiverio con flancos y muslos superiores ocres (Nardelli, 1993), se considera actualmente como posible aberración de plumaje.Machos
A diferencia de las hembras, que presentan variaciones cromáticas drásticas entre subespecies, los machos de Crax fasciolata son morfológicamente muy similares. Sin embargo, existen diferencias clave en tamaño y tonalidad que permiten distinguirlos: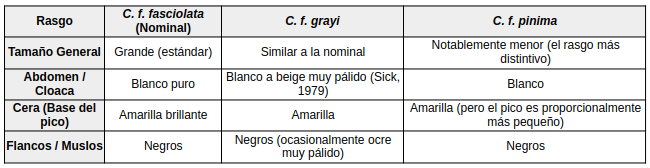
Diferencias Morfológicas entre Machos de la especie Hembras
Las hembras de Crax fasciolata son polimórficas, lo que significa que el plumaje varía drásticamente según la subespecie. A diferencia de los machos (casi todos negros), las hembras presentan patrones de barrado que son fundamentales para la identificación taxonómica.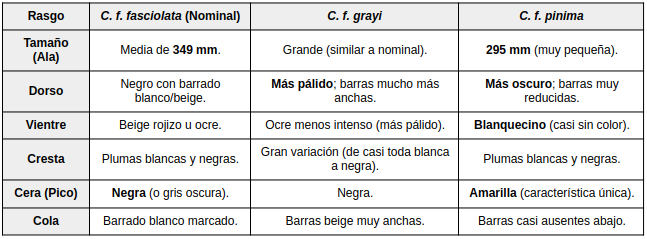
Diferencias Morfológicas entre Hembras de la especie Detalles Específicos para Identificación
• La Cera Amarilla: Es el rasgo más fácil para identificar a la hembra de C. f. pinima en campo, ya que las otras dos subespecies tienen la base del pico oscura.
• El ala de grayi: Esta subespecie se distingue por una mancha ocre grande y conspicua en el borde del ala (coberteras externas) que no presenta barrado, a diferencia de la nominal donde el barrado es continuo (Vaurie, 1968).
• Patrón Clinal: Existe una transición gradual; las aves son más oscuras en el noreste (pinima) y se vuelven más claras y con barras más anchas hacia el suroeste (grayi).El «Caso xavieri«: Como mencionamos, en los textos académicos existió una propuesta de subespecie (C. f. xavieri) basada en un macho que tenía flancos y muslos de color ocre. Hoy se sabe que esto no es una característica de una población, sino una aberración individual de plumaje (Nardelli, 1993).
La cresta: En las tres subespecies, el macho presenta una cresta de plumas rizadas totalmente negras. No presentan las plumas blancas que suelen verse en la cresta de las hembras.
Piel orbital: Todos presentan una zona de piel desnuda negra alrededor del ojo, pero en C. f. pinima esta área puede verse reducida debido al menor tamaño de la cabeza.
Aunque los machos de pinima se parecen mucho a los nominales, el hecho de que sean significativamente más pequeños es uno de los argumentos técnicos (Tobias et al., 2010) que refuerza su separación como especie independiente.Variación geográfica
Se ha documentado variación en el tamaño corporal y, especialmente, en el plumaje de las hembras, mientras que los machos muestran menor variabilidad. Las diferencias incluyen el color de fondo, la extensión del barrado ventral y dorsal, la intensidad de la coloración del dorso y el aspecto de la cresta. Esta variación presenta un patrón clinal general, desde poblaciones más oscuras en el noreste hasta formas más claras en el suroeste (Vaurie, 1967:1). No obstante, la magnitud de dicha variación se ve influida por una elevada variabilidad individual (Teixeira & Sick, 1986).Relaciones filogenéticas
Los análisis genéticos indican que Crax fasciolata y el pavón guayanés (Crax alector) constituyen especies hermanas (Pereira & Baker, 2004:689). Asimismo, la variabilidad genética intraespecífica de C. fasciolata ha sido evaluada en estudios previos (Pereira & Wajntal, 2001:301). - Crax fasciolata fasciolata (Spix, 1825) (Especie nominal)
-
Muitú (Crax fasciolata)

Spix (1781-1826). Avium species novae, quas in itinere per Brasiliam annis MDCCCXVII-MDCCCXX. Lámina LXII El Crax fasciolata se distribuye desde la Amazonia oriental hacia el sur, a través del centro de Brasil, hasta Paraguay y el norte de Argentina. Busca alimento en el suelo, en bosques y sus linderos, donde es bastante común. Al igual que otros del género Crax, esta especie presenta dimorfismo sexual: el macho es mayormente negro con la cloaca blanca, mientras que la hembra tiene el dorso rayado de negro y la parte inferior de color naranja a rojizo. Es el único con una extensa piel negra desnuda en la cara (Kirwan et al., 2025).
Nota antropológica: (Cebolla Badie, 2000:72) «Moitú (Crax fasciolata). Los indígenas de más edad recordaban que en el pasado lo cazaban y consumían su carne pero hace mucho tiempo que no ven un ejemplar. Explicaron que la hembra de la especie es batará y que al Mytu lo llaman jaku ramingua por su parecido con los jaku. Esta ave está casi extinta y es llamativo que los paisanos que los paisanos la asocien con los jaku porque efectivamente pertenecen todos al grupo Cracidae» (En Cadogan, mytu yapy katy ojeaity apo, el mytu anida cerca del naciente).
Nombres comunes: Faisán criollo, Pava de monte, Pavo de monte, Pavo real del monte, Pavón muitú, Mitú, Mitú-yacú, Moitú, Muitú común, Mutú. (de la Peña, 2020:287)
En otros idiomas
Breves apuntes históricos
- Avium species novae, quas in itinere per Brasiliam annis MDCCCXVII-MDCCCXX. Spix, Johann Baptist von, 1781-1826, Schmidt, Matthias. Monachii, Typis Franc. Seraph. Hübschmanni, 1824-1825: pp 48; Lámina 62a (Crax fasciolata)
- Apuntamientos para la historia natural de los páxaros del Paragüay y Rio de la Plata. (1805) de Azara, F. Imprenta de la Viuda de Ibarra. Vol. 3:83. (Mitú)
- Zur Ornithologie Brasiliens: Resultate von Johann Natterers Reisen in den Jahren 1817 bis 1835, von Pelzeln, 1870. Abtheilung 3: 287/341. (Crax fasciolata pinima)