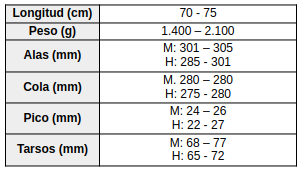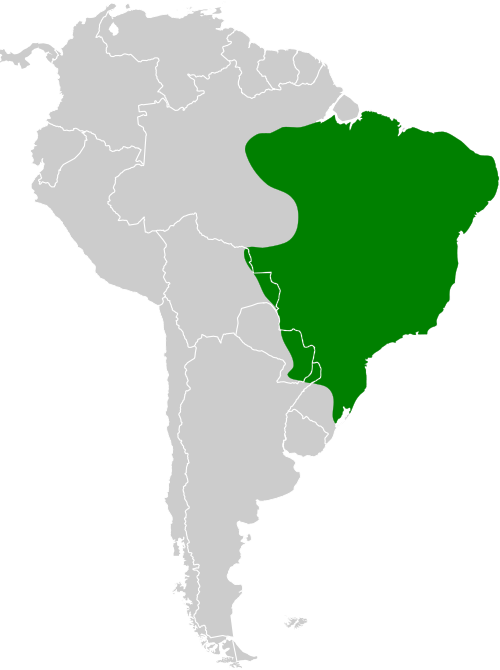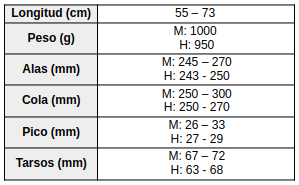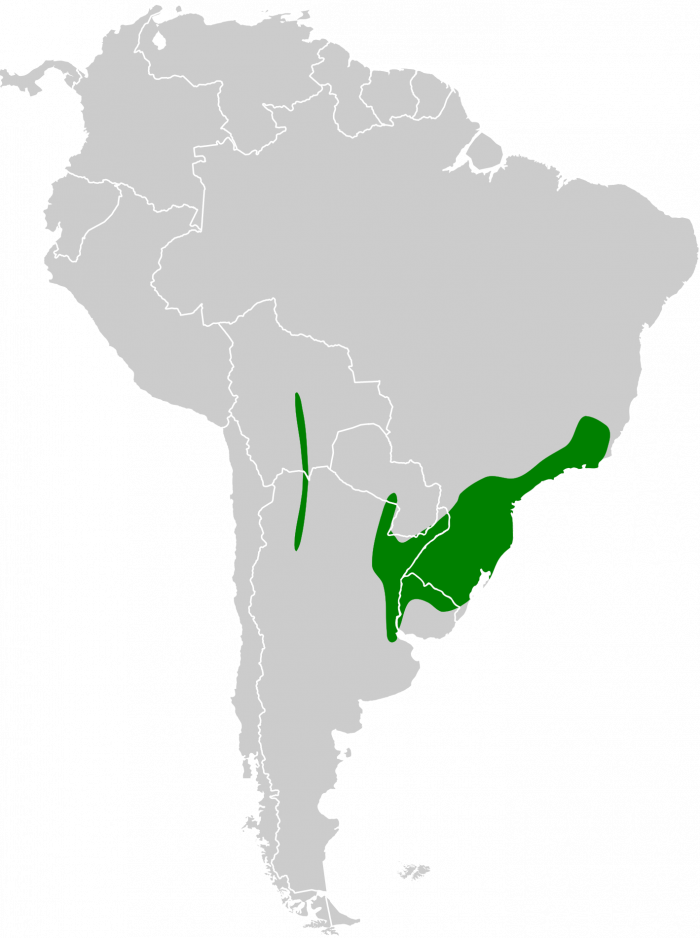
Especie de amplia distribución en el este y sur de América del Sur. En Brasil se encuentra principalmente desde los estados de Espírito Santo, Río de Janeiro y el sureste de Minas Gerais, hacia el sur a través de São Paulo y Paraná hasta Santa Catarina (Vaurie, 1966:18; Vaurie, 1968:198), con registros adicionales en el sur de Bahía (Souza, 1999:6). También ha sido registrada en el sureste de Paraguay (Smith & Derna, 2015:213), Uruguay y el noreste de Argentina, incluyendo las provincias de Buenos Aires, Entre Ríos, Corrientes, Misiones, el Chaco oriental y el norte de Santa Fe (Vaurie, 1968:198; del Hoyo & Motis, 2004:379; del Hoyo & Kirwan, 2024). Otros autores amplían su distribución a Bolivia (Vaurie, 1968:198; Sick, 1993:196; Stotz et al., 1996:303; de la Peña, 2025:277).
La brecha entre las áreas de distribución probablemente representa una modificación secundaria de la distribución causada por un aumento de la aridez en el Chaco, que obligó a la especie a expandirse hacia el oeste.
HÁBITAT
habita principalmente ambientes forestales, incluyendo bosques húmedos, selvas, bosques de galería y formaciones ribereñas asociadas a ríos, riachos y arroyos (Contreras et al., 2014:167; de la Peña, 2025:276). También se registra en bosques tropicales y subtropicales, bosques siempreverdes y sectores del Bosque Atlántico (Stotz et al., 1996:148). Las poblaciones del sur muestran una estrecha asociación con los bosques de galería, aunque la especie también utiliza islas fluviales con vegetación secundaria y plantaciones maduras rodeadas de pastizales, parches boscosos y áreas de regeneración secundaria (Malzof et al., 2012:7; del Hoyo & Motis, 2004:380).
Se distribuye desde las tierras bajas hasta zonas montanas, ocupando tanto el estrato medio como el suelo y la canopia del bosque (Stotz et al., 1996:148). Ha sido registrada desde el nivel del mar hasta aproximadamente 2.300 msnm, especialmente en regiones donde las áreas bajas son demasiado secas para mantener bosques siempreverdes o donde la cobertura forestal original ha sido removida (del Hoyo & Motis, 2004:380).
En Brasil, la especie es considerada residente común en parches de bosque, vegetación secundaria, matorrales, tierras agrícolas y bosques de galería (Belton, 1984:480). En Rio Grande do Sul ocupa gran parte del estado, aunque resulta menos frecuente en la franja costera, donde solo existen registros aislados. Las observaciones disponibles indican una mayor presencia en áreas boscosas y serranas vinculadas a las colinas del sur. En Itatiaia y Serra dos Órgãos en Río de Janeiro, habita bosques altos y montanos, donde con frecuencia constituye la única especie del género presente (Sick, 1997:277).
Las poblaciones septentrionales pueden encontrarse en bosques de Araucaria, aunque muestran preferencia por el Bosque Atlántico (Guix, 1997:201). En contraste, la especie puede resultar escasa en los bosques más densos, posiblemente debido a la mayor abundancia de depredadores mamíferos y a sus hábitos de alimentación terrestre (del Hoyo & Motis, 2004:381). Asimismo, ocasionalmente incursiona en matorrales y áreas agrícolas, incluidas plantaciones maduras (Malzof et al., 2012:7; del Hoyo & Motis, 2004:380).
De hábitos mayoritariamente sedentarios, esta especie presenta, no obstante, desplazamientos altitudinales estacionales en el sur de São Paulo, al sureste de Brasil. La evidencia disponible sugiere que algunos individuos descienden entre noviembre y febrero hacia zonas de menor elevación, probablemente con el fin de aprovechar la fructificación de numerosas especies de Myrtaceae presentes en dichos ambientes (del Hoyo & Motis, 2004:379).
Ecorregiones: Selva Paranaense, Delta e Islas del Paraná, Chaco húmedo, Campos y malezales
DISTRIBUCIÓN GEOGRÁFICA EN ARGENTINA
Según eBird, en el año 2026 existen 2580 observaciones, 528 fotografías y 16 registros sonoros de la especie en Argentina. Históricamente, Penelope obscura fue citada para las provincias de Formosa, Chaco, Misiones, Corrientes, noreste de Santa Fe, este de Entre Ríos y el delta del Paraná (de la Peña, 2015:190). Registros similares incluyen además el noreste de Santiago del Estero (Chébez, 2009:183; de la Peña, 2025: 277; de la Peña, 2015:190).
No obstante, la distribución actual parece considerablemente más restringida. En Argentina, las poblaciones relictuales confirmadas se localizarían principalmente en el delta inferior del Paraná y su área de influencia, en la región de Curuzú Cuatiá (Corrientes) y en los parques provinciales Urugua-í y Profundidad, en Misiones (del Hoyo & Motis, 2004:379). La población más importante se encontraría confinada al sector oriental del delta del Paraná, en el noreste de Buenos Aires (Césari & Domínguez Alonso, 1974; Pearman in prep.). Asimismo, se considera escasa en el sudeste de Entre Ríos, donde se registró una hembra en el Parque Nacional El Palmar, y en el sur-centro de Corrientes, particularmente en la estancia El Remanso y en las cuencas de los ríos Curuzú Cuatiá y Miriñay.
Diversos registros históricos para otras provincias han sido cuestionados por carecer de documentación adecuada o confirmación posterior. Los reportes de Chaco y Formosa son considerados provisionales debido a la ausencia de evidencias consistentes y a la falta de registros confirmados en el Chaco argentino y paraguayo. De manera similar, las menciones para el noreste de Santiago del Estero han sido calificadas como dudosas (Nores e Yzurieta, 1988; Chébez, 1994). La situación en el noreste de Santa Fe también permanece incierta, ya que los registros disponibles son escasos, antiguos o insuficientemente documentados, por lo que requieren verificación adicional.
Actualmente se citan registros en las provincias de Buenos Aires, Chaco, Formosa, Entre Ríos, Corrientes y Misiones. (Citas y observaciones)