InfoNatura: Animals and Ecosystems of Latin America 2007
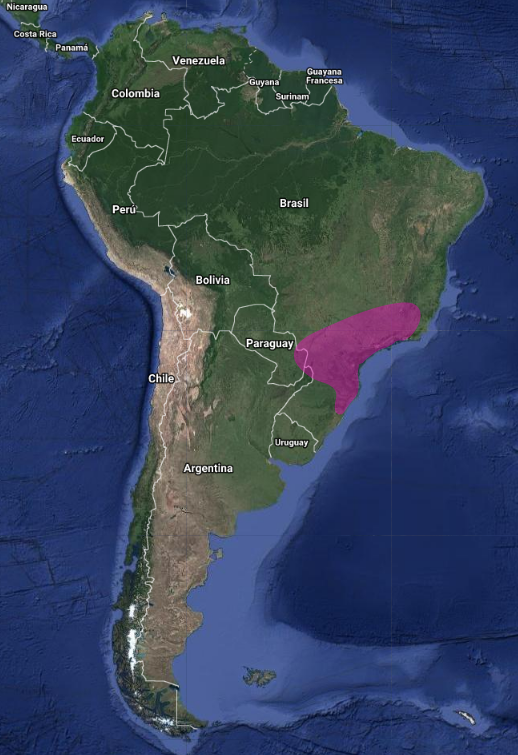
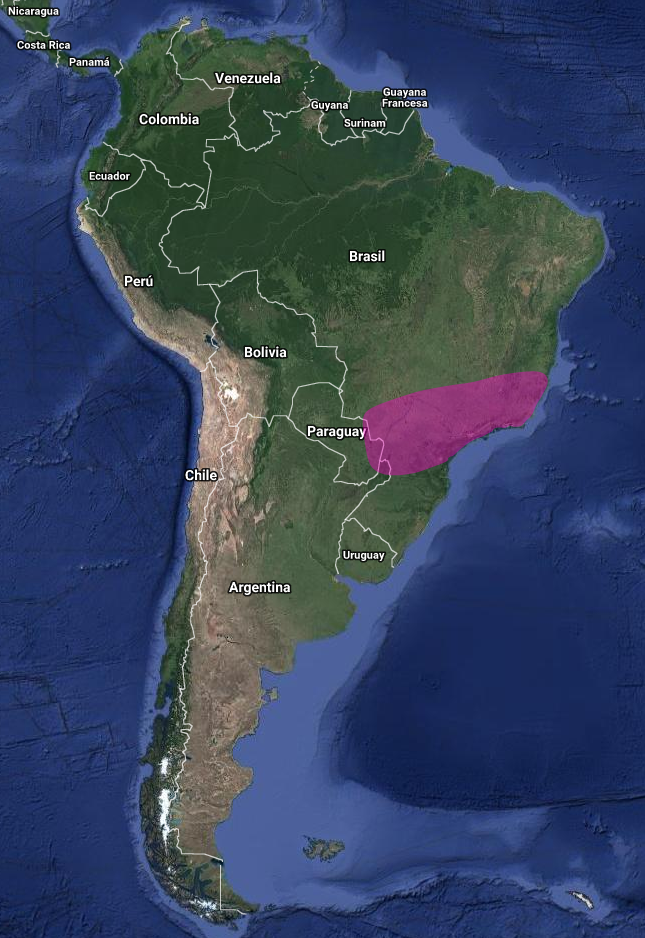
En América del Sur tiene una distribución amplia, pero dispersa, con registros en Este de Paraguay, Sureste de Brasil y Noreste de Argentina (Misiones) (Olrog, 1979:138; Sick, 1993:318; Brooks et al., 1993:118; Brooks et al., 1995:131; Chantler, 2000:108; Narosky & Yzurieta, 2010:197; Pearman et al., 2010:352; Stotz et al., 1996:317; Biancalana, 2015:402-403).
En Brasil, se puede encontrar en Rio Grande do Sul, Paraná, São Paulo, Río de Janeiro, Minas Gerais, Goiás, Tocantins, Bahía y Ceará (Belton, 1984:564; Pacheco et al., 1996:5; Sick, 1997:497; Barnett et al., 2004:54; Silveira et al., 2005:52; Straube et al., 2005:22; Vasconcelos et al., 2006:74; Stopiglia & Raposo, 2007:52; Albano & Girão, 2008:145; Pearman et al., 2010:351; Biancalana et al., 2012:1; Biancalana, 2015:402).
Se ha confirmado la presencia de nidos activos en Tocantins, Minas Gerais, Río de Janeiro y Paraná en Brasil (Vasconcelos et al., 2006:74; Biancalana et al., 2012:1) y en Misiones, Argentina (Pearman et al., 2010:352).
Brooks et al. (1995:131) comentan que en Paraguay, Bertoni (1939:26) reportó la especie para «Paraná», pero sin evidencias y Hayes (1995:129) consideró este informe como «hipotético». Sin embargo, comunican nuevos registros que pueden indicar que la especie es un residente escaso en el este de Paraguay, con un aumento en el número de individuos debido a la llegada de migrantes en la primavera austral, que han pasado el invierno más al norte.
Al menos en la porción sur de su rango, puede estar presente solo de manera estacional, por ejemplo, principalmente de septiembre a marzo en Río Grande do Sul, Brasil (Belton, 1984:564) y de agosto a marzo en Misiones, Argentina (Pearman et al., 2010:352). Aunque se ha informado de su presencia en Bolivia (Chantler, 2000:108), no hay registros confirmados en ese país (Pearman et al., 2010:356).
Distribución geográfica en Argentina: En la provincia de Misiones (Citas y observaciones)
HÁBITAT
Se lo observa sobre cañones, pantanos, bosques montanos húmedos, bosques templados y áreas de crecimiento secundario (Belton, 1984:564; Stotz et al., 1996:174; Vasconcelos et al., 2006:74; Di Giácomo et al., 2007:494).
Frecuenta selvas entre los 700 y los 2100 m de altura (de la Peña, 2015:337; de la Peña, 2020:254; Stotz et al., 1996:174; Hernández, 2020)
La especie tiene una fuerte asociación con la Mata Atlántica, con registros dispersos en áreas de transición entre otros biomas (Barnett & Pearman, 2001:48; Vasconcelos et al., 2006:74; Stopiglia & Raposo, 2007:52; Albano & Girão, 2008:145; Biancalana et al., 2012:2; Biancalana, 2015:403).
Son aves algo escasas y poco conocidas entre nosotros, debido especialmente a la extremada velocidad del vuelo, lo que hace muy difícil cazarlas. Nunca se posan sobre las ramas de los árboles, pero trepan con facilidad sobre las superficies verticales de las rocas ayudándose con sus largas y robustas uñas y sirviéndose de los mástiles rígidos de las plumas de la cola como punto de apoyo a la manera de los carpinteros. Se posan «en acantilados y detrás de cascadas» (Belton, 1984:564). Los despliegues de cortejo, si existen, aparentemente no han sido descritos, pero se presume que son al menos socialmente monógamos.
Forrajean en pequeños grupos de tres a seis individuos (Sick, 1993:318; de la Peña, 2015:337), aunque Belton (1984:564) encontró un aparente dormidero de 100 o más individuos. A veces junto a otras especies de vencejos, como el vencejo de collar (Streptoprocne zonaris), el vencejo de nuca blanca (Streptoprocne biscutata) y el vencejo chico (Chaetura cinereiventris) (Belton, 1984:564; de la Peña, 2020:253).
Migraciones: No existen evidencias claras de hábitos migratorios en esta especie, aunque ha sido considerada como tal en la literatura. Las deducciones sobre migración se basan solamente en la falta de registros en los meses de invierno, lo que hace necesario futuros estudios (Stopiglia & Raposo, 2007:54).
La ausencia total de registros confirmados en Argentina entre mediados de Marzo y comienzos de Agosto, sugiere dispersión post-reproductiva o migración austral. Las observaciones sugieren que, puede ser parcialmente migratoria, con afluencia de aves en primavera (Stotz et al., 1996:414; Pearman et al., 2010:356).
Vocalizaciones: Llamadas de vuelo rápidas “ti-i-i-i-a-a-a” o “Tiju-Tiju”. También un corto y seco “chit” o “jit”, a menudo repetido.
.
ALIMENTACIÓN
insectívoro, pero la dieta no se conoce con detalle (Hernández, 2020; de la Peña, 2020:254; Bodrati & Salvador, 2015:80).
REPRODUCCIÓN
Se reproducen a partir de febrero en Tocantins (Biancalana et al., 2012:2), de diciembre a enero en Minais Gerais (Vasconcelos et al., 2006:74), de noviembre a febrero en Paraná (Pichorim et al., citado por Vasconcelos et al. 2006:75), y noviembre-enero en Misiones (Pearman et al., 2010:358). También se reporta reproducción entre septiembre y marzo en Río de Janeiro (Fichberg et al., citado por Vasconcelos et al., 2006:75); los especímenes colectados en Rio Grande do Sul en octubre y noviembre estaban en condiciones de reproducción (Belton, 1984:564). En Argentina han sido localizados nidos entre noviembre y febrero en la provincia de Misiones (Pearman et al., 2010:353-355; de la Peña, 2020:254).
Los nidos tienen estructura en forma de copa o media luna compuesta por tallos de helecho, raicillas, musgo, hojas, tierra, guijarros y barro (Vasconcelos et al., 2006:74; Stopiglia & Raposo, 2007:55; Pearman et al., 2010:356; Biancalana et al., 2012:3; Biancalana, 2015:404). Destaca la ausencia de saliva en su construcción (Sick, 1993:318; Stopiglia & Raposo, 2007:49; Biancalana, 2015:407).
La mayoría de los nidos se encuentran en repisas naturales en las paredes de los acantilados, pero algunos están unidos directamente a la pared. La altura desde el suelo al nido, en la pared del acantilado, difiere mucho, oscilando entre 1,5 y 60 m (Vasconcelos et al., 2006:74; Pearman et al., 2010:357; Biancalana et al., 2012:3). Estas aves tienden a anidar lo más cerca posible de las cascadas sin mojar el nido, ya sea que la cascada esté en una cueva pequeña o en una cueva grande en un río (Pearman et al. 2010:357). Biancalana (2015:407), comenta «Vasconcelos et al. (2006:74) informaron que los nidos de «Sooty Swift» están protegidos del rocío del agua, lo cual difiere de las observaciones de este estudio; los nidos estaban expuestos a goteo y rocío de agua, y los polluelos presentaban plumas con gotas de agua. Una situación similar fue observada por Pearman et al. (2010:357) en Argentina, donde uno de los nidos de «Sooty Swift» estaba en contacto permanente con el rocío de la cascada».
La puesta es de un solo huevo blanco (Vasconcelos et al., 2006:74; Pearman et al., 2010:357; Biancalana, 2012:3). Ambos adultos comparten la incubación durante la noche (Biancalana, 2015:404).
Los polluelos muestran ojos cerrados y ausencia de plumaje en los primeros días de vida, crecimiento de plumaje gris oscuro durante las primeras dos semanas y la presencia de plumas con puntas blancas (Biancalana et al., 2012:4).
Los adultos abandonan el nido y pasan grandes periodos del día buscando insectos. Se los ha observado alimentando a sus polluelos por la noche, lo que probablemente se relaciona con la alta actividad de insectos durante el final de la tarde y los períodos nocturnos (Biancalana et al., 2015:408).
Nota: Las descripciones del nido y los huevos del vencejo ahumado chico realizadas por Dabbene (1918:193) se refieren al vencejo parduzco (Cypseloides rothschildi), no al vencejo negruzco.
No dimorfismo sexual. Tamaño mediano.
Plumaje pardo negruzco, compacto y cerrado al cuerpo.
La cabeza marrón con un parche negro rodeando los ojos y y frente y auriculares ligeramente más claras. Iris marrón. Pico negro
El dorso es marrón oscuro, más oscuro que la cabeza y la nuca, y un poco más oscuro que la rabadilla.
Alas más bien largas. Las cubiertas alares y zona exterior de remeras primarias; la superficie inferior es más grisácea. La zona interna de las remeras primarias, secundarias y cubiertas alares son marrón más pálido. Ribeteado pálido muy indistinto en las cubiertas y remeras.
Cola corta y cuadrada. La parte superior y superficial de las timoneras marrón-negruzco; en la zona más inferior, grisácea.
Tarsos y dedos de los pies negros. (Belton, 1984:564; Chantler, 2000:108; de la Peña, 2015:337; Hernández, 2020; de la Peña, 2020:253)
Medidas: (Eisenmann & Lehmann, 1962:8; Belton, 1984:564; Sick, 1993:318; Sick, 1997:429; Chantler, 2000:108; Roesler et al., 2009:226; de la Peña, 2015:337; de la Peña, 2020:253; del Hoyo, 2020:128).
Descrita originalmente como «Hemiprocne fumigatus» Streubel 1848, con localidad tipo en Paraguay y Brasil (restringida a Curitiba, Paraná, Brasil por Zimmer, 1945:587), el «Cypseloides fumigatus» ha sido objeto de debate taxonómico. Algunos autores, como Zimmer (1945:589), han clasificado al «Cypseloides rothschildi» (Vencejo de Rothschild) como una subespecie de «fumigatus», mientras que la mayoría sigue a Rogers (1939) en reconocer a «fumigatus» y «rothschildi» como especies separadas. Las relaciones filogenéticas dentro del género «Cypseloides» no han sido completamente resueltas. Short (1975:238), consideró que «fumigatus» y «rothschildi» forman una superespecie junto con «Cypseloides niger» (Vencejo negro) y «Cypseloides lemosi» (Vencejo pechiblanco).
Short (1975:238), señaló que considera a «niger» (distribuido en la región Neártica, las Indias Occidentales y América Central), «lemosi» (de Colombia), «rothschildi» (del noroeste de Argentina: Salta, Santiago del Estero, Tucumán) y «fumigatus» (del sureste de Brasil: Espirito Santo hasta Rio Grande do Sul) como especies que comprenden una superespecie (Hernández, 2020).
El vencejo negruzco (Cypseloides fumigatus) es una de las aves menos estudiadas, principalmente debido a las dificultades de identificación en campo y a sus hábitos esencialmente aéreos. Normalmente se la encuentra volando sobre bosques montanos o tropicales de tierras bajas, en selvas siempreverdes y matorrales de crecimiento secundario. De tamaño medio dentro del género, presenta una cola corta y cuadrada, con alas relativamente largas en comparación con el cuerpo. Su plumaje es marrón negruzco, con el manto más oscuro que la cabeza. No presenta dimorfismo sexual, lo que a menudo lleva a confundirla con «Cypseloides senex» (Stopiglia & Raposo, 2007:49). Se distribuye principalmente en el Este de Brasil, incluyendo también el Este de Paraguay y Noreste de Argentina (Hernández, 2020).
Nombres comunes:Vencejo negruzco(Argentina, Paraguay), Vencejo pardo, Vencejo parduzco, Vencejillo negro, Vencejo fuliginoso, Vencejo pardo chico, Vencejo panza gris. En Brasil: Andorinha-das-tormentas, andorinhão-preto-da-cascata, andorihão-cinzento, taperuçu-cinza, taperucu-preto, Taperuçu-preto; En guaraní: «Mbyju’i hû» (Chebez, 1996: 135; de la Peña, 2015:337; de la Peña, 2020:253)
Orden Caprimulgiformes: 8 familias. Con una vasta distribución mundial, que abarca 112 especies y 19 géneros (Winkler et al, 2020; del Hoyo, 2020:128-130), los vencejos están bien representados en la región neotropical, donde también se encuentran las especies de mayor tamaño (Streptoprocne); en Argentina se registran 9 especies y 6 en la provincia de Misiones.
Nota taxonómica: Tradicionalmente, los vencejos se han clasificado dentro del orden Apodiformes; sin embargo, aquí se incluyen en el orden Caprimulgiformes basándonos en estudios filogenéticos recientes (Barrowclough et al., 2006; Ericson et al., 2006; Hackett et al., 2008:1765; Braun y Huddleston, 2009; Mayr, 2010:126; Mayr, 2011:69). Aunque algunas divisiones dentro de este grupo permanecen controvertidas, existe un consenso general en que los vencejos están más estrechamente relacionados con los vencejos arborícolas del sudeste asiático (Familia Hemiprocnidae). Estas dos familias, a su vez, son grupos hermanos de los colibríes. Las tres familias, que en conjunto constituyen los Apodiformes tradicionales, son hermanas de los atajacaminos (Barrowclough et al., 2006; Ericson et al., 2006; Hackett et al., 2008:1765; Braun y Huddleston, 2009:959; Mayr, 2010:126; Mayr, 2011:69). Esta última relación de grupo hermano con una familia caprimulgiforme tradicional es uno de los factores que motivó la decisión de incluir a los Apodiformes tradicionales dentro de los Caprimulgiformes.
La evidencia actual indica que estas familias pertenecen a un único clado, Caprimulgiformes + Apodiformes (Ericson et al., 2006; Hackett et al., 2008). Todos los árboles filogenéticos apoyan un clado que incluye a las aves nocturnas (los “caprimulgiformes” tradicionales, excluyendo a los búhos) y las aves apodiformes (vencejos y colibríes; figura 1, nodo 1) (Mayr, 2002; Cracraft et al., 2004) y, por primera vez, sugieren la monofilia de un clado que incluye a todos los taxones tradicionalmente ubicados en “Caprimulgiformes” y Apodiformes. La topología obtenida sugiere que los Apodiformes diurnos evolucionaron dentro de una radiación de aves nocturnas, indicando un ancestro nocturno para los Apodiformes (Ericson et al., 2006; Hackett et al., 2008).
El orden Caprimulgiformes comprende cinco familias de aves adaptadas a la actividad nocturna. Este orden ha sido considerado monofilético, pero evidencia reciente sugiere que los vencejos y los colibríes (Apodiformes) pertenecen a él. Para explorar la filogenia del grupo, se obtuvieron más de 2000 pb de secuencia de ADN de los genes citocromo b y c-myc para 35 taxones, representando todas las principales líneas evolutivas y grupos externos. Las secuencias no codificantes del gen c-myc no estaban saturadas, eran fácilmente alineables y contenían numerosas inserciones y deleciones informativas, señalando una utilidad amplia para la filogenia a nivel superior. La monofilia de cada una de las cinco familias tradicionales fue fuertemente confirmada, pero las relaciones entre familias fueron apenas resueltas. La estructura del árbol argumenta en contra del estatus de familia para Eurostopodus y Batrachostomus, que deberían ser retenidos en Caprimulgidae y Podargidae, respectivamente. El género Caprimulgus y ambas subfamilias de Caprimulgidae parecen ser polifiléticos. La filogenia elucida la evolución de rasgos adaptativos como la nocturnidad y la hipotermia, pero sigue siendo una cuestión abierta si la nocturnidad evolucionó una o múltiples veces (Braun & Huddleston, 2009:957).
COMPORTAMIENTO
Cosmopolitas y gregarios, los vencejos presentan dimorfismo sexual mínimo (Sick, 1997:423). Estas aves han sido frecuentemente confundidas con las golondrinas debido a su apariencia similar, sin embargo, este parecido es meramente superficial; sus características anatómicas las sitúan más cerca de los colibríes. Los vencejos son insectívoros y eminentemente aéreas; nunca se posan sobre ramas de árboles, pero trepan con facilidad superficies verticales de rocas, ayudándose con sus largas y robustas uñas y utilizando los mástiles rígidos de sus plumas caudales como apoyo, de manera similar a los carpinteros (Dabbene, 1917:3).
Viven en continuo movimiento y se cuentan entre las aves más dinámicas del planeta, con una notable capacidad para reducir abruptamente sus velocidades (Sick, 1997:423). Su vuelo es permanente, rápido y sinuoso, a menudo a gran altura, y se alimentan en vuelo. Sin embargo, durante condiciones de tiempo lluvioso y frío, permanecen todo el día en el nido.
A menudo confundidos con las golondrinas debido a su cuello corto y pico de base ancha que les facilita cazar insectos en vuelo, los vencejos tienen patas pequeñas y cortas, uñas fuertes, curvas y afiladas, y un hallux relativamente corto. Ponen varios huevos blancos en nidos situados en huecos de árboles, grietas en rocas o debajo de cascadas. Su pico es corto y arqueado, con una boca grande, mientras que sus alas son largas, estrechas y curvadas, sobrepasando la punta de la cola cuando están plegadas. El tarso es generalmente corto y a veces emplumado, al igual que los dedos.
Las alas de los vencejos son extremadamente largas y estrechas debido a la cortedad de las rémiges secundarias; sobrepasan la extremidad de la cola cuando están plegadas y adoptan una forma de hoz al estar extendidas. Los mástiles de las rectrices, siempre 10 en número, son rígidos y en algunos casos (Chaetura, Streptoprocne) están más o menos desprovistos de barbas en la extremidad. El plumaje es compacto y ajustado al cuerpo, con una coloración general de un pardo uniforme o con algunas partes blancas o blancuzcas sucias (Dabbene, 1917:4).
Los vencejos se posan verticalmente en superficies ásperas, utilizando sus fuertes uñas, apoyando el cuerpo en callosidades del metatarso, una adaptación para mantenerse sobre superficies cortantes, y sobre su rígida cola. Este comportamiento provoca un gran desgaste en el plumaje, especialmente en las timoneras y rémiges primarias externas. Las «púas» terminales de las timoneras pueden desgastarse por completo, lo que es una señal segura de que el individuo está reproduciéndose y pasa mucho tiempo sobre la roca áspera alrededor del nido (Sick, 1997:422). Sus glándulas salivares, que se desarrollan más durante la reproducción, les permiten confeccionar el nido, reduciendo la función digestiva de la saliva en estos casos (Sick, 1997:422).
Poseen una adaptación rara en las aves: tienen la capacidad de caer en «sopor», adquiriendo la inmovilidad completa (Sick, 1997:422).
De vida aérea, se alimentan de la abundante variedad de insectos presentes en el cielo. Algunas especies están tan adaptadas a sus actividades aéreas que no pueden despegar desde terrenos llanos, y algunas regresan a la tierra únicamente para reproducirse. Utilizando barro o saliva adhesiva, los vencejos fijan sus nidos (o huevos) a plantas, paredes o acantilados. Estos métodos de construcción de nidos permiten un descenso pronunciado para el despegue y, al mismo tiempo, brindan protección contra los depredadores. Al igual que algunos de sus parientes caprimulgiformes, ciertos vencejos han desarrollado la ecolocalización para habitar en cavernas profundas, mientras que otros sobreviven a periodos de frío sin alimento entrando en estado de letargo.
Vocalizaciones: Las vocalizaciones de los vencejos son simples, mientras que su canto consiste en una serie de sonidos algo más elaborados (Sick, 1997:423). El género Streptoprocne emite vocalizaciones corales al atardecer. En el caso de Chaetura, especialmente C. andrei, cuando están inquietos en su aterrizaje cerca del nido o al agruparse en bandadas, producen fuertes golpes con las alas contra el sustrato de manera amenazante. Este ruido es distinto del que producen las palomas al batir una ala contra la otra.
HÁBITAT
Habitan en una amplia variedad de hábitats terrestres, pero viven predominantemente en el aire, desplazándose con las masas de aire y las corrientes de convección locales para aprovechar las concentraciones locales de presas. Al depender de presas aéreas, a menudo se alejan considerablemente de sus nidos mientras buscan alimento para sí mismos y para sus crías.
ALIMENTACIÓN
Se alimentan casi exclusivamente de insectos voladores y otros artrópodos aéreos.
REPRODUCCIÓN
Los vencejos son aves monógamas que presentan cuidados biparentales. Las ubicaciones de los nidos varían considerablemente entre las diferentes especies de vencejos: algunas anidan en cuevas oscuras, otras detrás de cascadas, otras debajo de los aleros de puentes y edificios, y otras en árboles huecos. Aunque la mayoría de los vencejos construyen sus nidos adheridos a una pared vertical con barro o saliva, esta práctica también puede variar. Los materiales utilizados en la construcción del nido incluyen musgos, pasto, palos, plumas, plantas y barro, frecuentemente unidos entre sí y al sustrato mediante saliva adhesiva.
El tamaño de la nidada varía de uno a cinco huevos alargados y blancos, y ambos padres comparten todas las tareas de cuidado. La incubación dura entre dos y cuatro semanas, y el período de cría se extiende de seis a diez semanas. Se ha observado que una especie pone dos nidadas de un solo huevo con 50 días de diferencia, siendo la segunda incubada por el polluelo en desarrollo de la primera. Muchas de las especies no migratorias mantienen vínculos de pareja a largo plazo, y al menos una especie se reproduce de forma cooperativa, con ayudantes provenientes de intentos de reproducción anteriores. (Winkler et al., 2020). Los polluelos nacen ciegos y abren los ojos al sexto día de vida. El periodo de nidificación coincide con la temporada de lluvias, cuando hay una mayor abundancia de insectos (Sick, 1997:424).
Durante el periodo de reproducción, los vencejos exhiben un modo diferente de volar; especies como Streptoprocne, Cypseloides y Chaetura, incluyendo Chaetura andrei, frecuentemente vuelan en grupos de tres, realizando elegantes giros con las alas casi inmóviles y arqueadas, intercalando a veces ritmos rápidos (Sick, 1997:424). Los Streptoprocne y Chaetura, que son más gregarios, se reúnen en grandes bandadas, volando en círculos mientras emiten sus llamadas. Se han observado ensayos de copulación en vuelo (Sick, 1997:424).
REFERENCIAS
Azara, F. de (1802). «Apuntamientos para la Historia Natural de los Páxaros del Paragüay y Rio de La Plata. Madrid MDCCCII (enlace)
Barrowclough, G.F., Groth, J.G. & L.A. Mertz (2006). The RAG-1 exon in the avian order Caprimulgiformes: phylogeny, heterozygosity, and base composition. Molecular Phylogenetics and Evolution, 41 (1): 238–248.
Braun, M.J. & C.J. Huddleston (2009). A molecular phylogenetic survey of caprimulgiform nightbirds illustrates the utility of non-coding sequences. Molecular Phylogenetics and Evolution, 53 (3): 948–960. (enlace)
Chantler, P.J. (1995) Identification of three Chaetura swifts; Band-rumped Swift Chaetura spinicauda, Grey-rumped Swift Chaetura cinereiventris and Pale-rumped Swift Chaetura egregia. Cotinga 4: 44–51. (enlace)
Dabbene, R. (1917) Los vencejos de la República Argentina. Hornero, 1 (1): 003-008. (enlace)
Dabbene, R. (1918) Nidos y huevos de vencejos. Hornero, 1 (03): 193-193. (enlace)
del Hoyo, J. ed. (2020). All the Birds of the World. Lynx Edicions. Barcelona, España. 967 pp. ISBN 978-84-16728-37-4
Ericson, P.G., Anderson, C.L., Britton, T., Elzanowski, A., Johansson, U.S., Kallersjo, M., Ohlson, J.I., Parsons, T.J., Zuccon, D. & G. Mayr (2006). Diversification of Neoaves: integration of molecular sequence data and fossils. Biology Letters, 2 (4): 543–547. (enlace)
Hackett, S.J., Kimball, R.T., Reddy, S., Bowie, R.C., Braun, E.L., Braun, M.J., Chojnowski, J.L., Cox, W.A., Han, K.L., Harshman, J., Huddleston, C.J., Marks, B.D., Miglia, K.J., Moore, W.S., Sheldon, F.H., Steadman, D.W., Witt, C.C. & T. Yuri (2008). A phylogenomic study of birds reveals their evolutionary history. Science, 320 (5884): 1763–1768. (enlace)
Mayr, G. (2010). Phylogenetic relationships of the paraphyletic ‘caprimulgiform’ birds (nightjars and allies). Journal of Zoological Systematics and Evolutionary Research, 48 (2): 126–137. (enlace)
Mayr, G. (2011). Metaves, Mirandornithes, Strisores and other novelties – a critical review of the higher-level phylogeny of neornithine birds. Journal of Zoological Systematics and Evolutionary Research, 49 (1): 58–76. (enlace)
Remsen, J.V., Jr., J.I. Areta, E. Bonaccorso, S. Claramunt, G. Del-Río, A. Jaramillo, D. F. Lane, M. B. Robbins, F. G. Stiles & K. J. Zimmer. Version [date]. A classification of the bird species of South America. Museum of Natural Science, Louisiana State University. http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm VERSION: 16 May 2024
Sick, H. (1997). Ornitologia Brasileira. Editora Nova Fronteira, Rio de Janeiro, Brazil. 912 pp. ISBN 85-209-0816-0
Winkler, D.W., S.M. Billerman, and I. J. Lovette (2020). Swifts (Apodidae), version 1.0. In Birds of the World (S. M. Billerman, B. K. Keeney, P. G. Rodewald, and T. S. Schulenberg, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.apodid1.01
Alvarenga, H.M.F., Höfling, E. & L.F. Silveira (2002). Notharchus swainsoni (Gray, 1846) (Bucconidae) é um espécie válida. Ararajuba, 10 (1): 73-77. (enlace)
Aleixo, A. & M. Galetti (1997). The conservation of the avifauna in a lowland Atlantic forest in south-east Brazil. Bird Conservation International, 7 (3): 235–261. (enlace)
ArgentAvis. (s.f.). Proyecto “Argentavis – Aves de Argentina” (enlace)
Barnett, J.M. & M. Pearman (2001). Lista comentada de las aves argentinas. Lynx Edicions, Barcelona. 164 pp. ISBN 84-87334-32-6
Bodrati, A. & K. Cockle (2007). Alta cuenca del Arroyo Alegría. En Di Giacomo, A. S., M. V. De Francesco y E. G. Coconier (editores). 2007. Áreas importantes para la conservación de las aves en Argentina. Sitios prioritarios para la conservación de la biodiversidad: 290-292. Temas de Naturaleza y Conservación 5. CD-ROM. Edición Revisada y Corregida. Aves Argentinas/ Asociación Ornitológica del Plata, Buenos Aires. (enlace)
Bodrati, A., K. Cockle, J.M. Segovia, I. Roesler, J.I. Areta & E. Jordan (2010). La avifauna del Parque Provincial Cruce Caballero, Provincia de Misiones, Argentina. Cotinga, 32: 41-64. (enlace)
Bodrati, A. & S.A. Salvador (2015). Termitas (Isoptera, Blattodea, Insecta) en la dieta de las aves argentinas. Nuestras Aves, 5: 77-89. (enlace)
Brooks, T.M., R. Barnes, L. Bartrina, S.H.M. Butchart, R.P. Clay, E.Z. Esquivel, N.I. Etcheverry, J.C. Lowen & J. Vincent (1993). Bird Surveys and Conservation in the Paraguayan Atlantic Forest: Project CANOPY ’92 Final Report. BirdLife Study Report 57. BirdLife International, Cambridge, UK. (enlace)
Chebez, J.C. (1996). Fauna misionera, catálogo sistemático y zoogeográfico de los vertebrados de la provincia de Misiones (Argentina). Monografía No 5. Págs. 320. L. O. L. A. Bs.Aires. 320 pp. ISBN 950-9725-20-X
Cockle, K.L., A. Bodrati, M. Lammertink, B. B. Bonaparte, C. Ferreira & F. Di Sallo (2016). Predators of bird nest in the Atlantic Forest of Argentina y Paraguay. Wilson Jour. Ornith., 128: 120-131. (enlace)
Cory, C.B. (1919). Catalogue of Birds of the Americas. Publications of the Field Museum of Natural History, Zoological Series, 13 (2): 608. (enlace)
de la Peña M.R. (1999). Aves Argentinas. Lista y Distribución. Ed. L.O.L.A. (Literature of Latin America). Monografía N.º 18; 244 pp. Buenos Aires. ISSN 0328-1620
de la Peña, M.R. (2015). Aves Argentinas. Incluye nidos y huevos. 1º Ed. Santa Fe: Ediciones UNL; CABA: EUDEBA; Vol 1; 496 pp. ISBN 978-987-657-986-5
de la Peña, M.R. (2020:29). Aves Argentinas. Descripción, Comportamiento, Reproducción y Distribución. (Actualización). Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie), Vol 7: 1-344. ISSN 0325-3856
del Hoyo, J. ed. (2020). All the Birds of the World. Lynx Edicions. Barcelona, España. 967 pp. ISBN 978-84-16728-37-4
Giraudo, A.R., Baldo, J.L. & R.R. Abramson (1993). Aves observadas en el sudeste, centro y este de Misiones (República Argentina), con la mención de especies nuevas o poco conocidas para la provincia. Nótulas Faunísticas, 49: 1-13. (enlace)
Hayes, F.E. (1995). Status, Distribution and Biogeography of the birds of Paraguay. Monographs in Field Ornithology 1. American Birding Association, Colorado Springs, CO, USA. 224 pp. ISBN: 1-878788-30-2 (enlace)
Lepage, D. (2003). Avibase-La base de datos World Bird. Ficha: Notharchus swainsoni; Recuperado en mayo 2024 de https://avibase.bsc-eoc.org/species.jsp?avibaseid=5D3D72E119BEFB1C
López-Lanús, B., P. Grilli, E. Coconier, A. Di Giacomo & R. Banchs (2008). Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas /AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina. (enlace)
Matthews, A. & P. Smith (2017). Breeding observations on Buff-bellied Puffbird Notharchus swainsoni (Piciformes: Bucconidae) at rancho Laguna Blanca, San Pedro Department, Paraguay. Revista Brasileira de Ornitologia, 25 (1): 20-23. (enlace)
Narosky, T. & J.C. Chebez (2002). Aves de Iguazú: guía de identificación. 1º Ed. Vázquez Mazzini Editores, Buenos Aires. 128 pp. ISBN 987-9132-04-1
Narosky, T. & D. Yzurieta (2010). Birds of Argentina and Uruguay: a field guide. Vázquez Mazzini Editores, Buenos Aires, Argentina. ISBN 950-99063-6-0
Oliveira, R.C. & O. Arasaki (2011). Novos registros de Notharchus swainsoni (Galbuliformes: Bucconidae) na região noroeste do Estado do Paraná e ampliação da área de ocorrência no estado. Atualidades Orn., 162: 6-7. (enlace)
Olrog, C.Ch. (1979). Nueva Lista de la Avifauna Argentina. Ópera Lilloana XXVII. Ministerio de Cultura y Educación. Fundación Miguel Lillo. Tucumán, Argentina. 324 pp. ISSN 0078-5245
Partridge, W.H. (1954). Estudio preliminar sobre una colección de aves de Misiones. Rev. Mus. Arg. Cs. Nat. Zool., 3: 85-153.
Pereyra, J.A. (1950). Avifauna argentina (contribución a la ornitología). El Hornero, 9 (2): 178–241. (enlace)
Rasmussen, P.C., N. Collar & G.M. Kirwan (2020). Buff-bellied Puffbird (Notharchus swainsoni), version 1.0. In Birds of the World (J. del Hoyo, A. Elliott, J. Sargatal, D. A. Christie, and E. de Juana, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.bubpuf1.01
Saibene, C.A., M. A. Castelino, N.R. Rey, J. Herrera & J. Calo (1996). Inventario de las Aves del Parque Nacional “Iguazú”, Misiones, Argentina. Literature of Latin America, Buenos Aires, Argentina. 68 pp. ISSN 0328-1620
Schmidtuz, C., Güller, R., Di Santo, H., Chévez, R., Tahmazian, R. & A. Quinteros (2001). Nuevos registros para Misiones (Argentina) de la Viudita Coluda (Muscipipra vetula) y el Chacurú Grande (Notharchus macrorhynchus). Nuestras Aves, 41: 4. (enlace)
Short, L.L. (1975). A zoogeographic analysis of the South American Chaco avifauna. Bulletin of the American Museum of Natural History, 154 (3): 165–352. (enlace)
Short, L.L. (1972). Systematics and Behavior of South American Flickers (Aves, Colaptes). Bulletin of the American Museum of Natural History, 149 (1), New York. 110 pp. (enlace)
S.I.B. Sistema de Información de Biodiversidad de la Administración de Parques Nacionales, Argentina. Ficha: Notharchus swainsoni. Recuperado en mayo 2024 de https://sib.gob.ar/especies/notharchus-swainsoni
Sick, H. (1993). Birds in Brazil. A Natural History. Princeton University Press, Princeton, NJ, USA. ISBN 0-691-08569-2 (enlace)
Silveira, L.F., P.F. Develey, J.F. Pacheco, & B.M. Whitney (2005). Avifauna of the Serra das Lontras–Javi montane complex, Bahia, Brazil. Cotinga, 24: 45–54. (enlace)
Stotz, D.F., Fitzpatrick, J.W., Parker, T.A., & D.K. Moskovits (1996). Neotropical Birds: Ecology and Conservation. University of Chicago Press, Chicago, IL, USA, and London, UK. 478 pp. ISBN 0-226-77629-8
Xeno-canto (2024). Ficha: Buco de Swainson · Notharchus swainsoni · (Gray, 1846). Recuperado en mayo 2024 de https://xeno-canto.org/species/Notharchus-swainsoni
ESTADO DE CONSERVACIÓN SEGÚN BIRDLIFE INTERNATIONAL: PREOCUPACIÓN MENOR
Especie de amplia distribución, no alcanza los umbrales de vulnerabilidad bajo este criterio ( extensión < 20.000 km² combinada con su declinación o fluctuación, calidad/extensión del hábitat, o tamaño de la población y pequeño número de localizaciones o fragmentación grave)
La tendencia de la población parece ser estable y tampoco alcanza los umbrales de vulnerabilidad bajo los criterios de tendencia de las poblaciones (> 30% declinación en 10 años o 3 generaciones)
El tamaño de la población no ha sido cuantificado, pero tampoco se cree que alcance los niveles de vulnerabilidad según criterio de tamaño de la población (<10.000 individuos maduros con una declinación continua en su número > 10% en 10 años o 3 generaciones)
Por estas razones se considera a esta especie como de “preocupación menor»
Rara en Argentina (Misiones), aunque regularmente se registra en el área de Iguazú.
Estado de conservación Vulnerable (VU): según AA y SAyDS, 2008 (Aves Argentinas y Secretaría de Ambiente y Desarrollo Sustentable). López-Lanús, B., P. Grilli, E. Coconier, A. Di Giacomo y R. Banchs. Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas /AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina. 1 ed. AOP: Buenos Aires, Argentina (2008:82) Vulnerable (VU): según MADS y AA, 2017 (Ministerio de Ambiente y Desarrollo Sustentable de la Nación y Aves Argentinas). Categorización de las Aves de la Argentina según su estado de conservación. Informe del Ministerio de Ambiente y Desarrollo Sustentable de la Nación y de Aves Argentinas. 146 p. 2017 Vulnerable (VU): según SAyDS, 2010 (Secretaría de Ambiente y Desarrollo Sustentable). Categorización de aves autóctonas de Argentina. Lista de aves no registradas, endémicas y exóticas de Argentina (Res 348). 2010 Preocupación Menor (LC): según UICN (API) (Unión Internacional para la Conservación de la Naturaleza). Lista Roja de la UICN publicada en https://www.iucnredlist.org. Actualización en tiempo real
InfoNatura: Animals and Ecosystems of Latin America 2007
Se distribuye en el Sureste de Brasil, desde Espírito Santo hasta Santa Catarina, y se extiende hacia el este hasta Bahía y Mato Grosso do Sul; hacia el oeste hasta el este de Paraguay (localmente rara y poco común en otras regiones, como el Bajo Chaco y Campos Cerrados, así como en el centro de Paraguay (Hayes, 1995:102) y Noreste de Argentina (Misiones) (Oliveira y Arasaki, 2011:6; Cory, 1919:392; Schubart et al., 1965:177; de la Peña, 2020:30; Matthews & Smith, 2017:20; Olrog, 1979:149; Pereyra, 1950:235; de la Peña, 2020:30).
Distribución geográfica en Argentina (de la Peña, 2020:30) Notharchus swainsoni (Gray, 1846) en Misiones. (Citas y observaciones)
Migraciones
Posible migrante de verano, de septiembre a marzo, a la Mata Atlántica en el estado brasileño de São Paulo; Residente en Iguazú, en el extremo Noreste argentino.
Hábitat
Esta especie se encuentra típicamente en hábitats de bosque húmedo de tierras bajas, incluyendo el Bosque Atlántico (Barnett & Pearman, 2001:51; de la Peña, 2015:372), así como en áreas de bosque secundario alto y talado. Su rango altitudinal se extiende hasta aproximadamente de 0-900 msnm (del Hoyo, 2020:315; Silveira et al., 2005:52) hasta los 1.300 msnm (Stotz et al., 1996:190). Se ha documentado su presencia en tierras bajas del Bosque Atlántico en el sureste de Brasil (Alexio & Galetti, 1997:251). Ecorregiones:Selva Paranaense
Exhibe una serie de comportamientos y hábitos particulares según lo documentado por varias fuentes (Narosky & Chebez, 2002:68; Narosky & Yzurieta, 2010:214; Pereyra, 1950:235). Se ha observado que suele posarse en ramas altas, mostrando un vuelo corto y algo torpe (Narosky & Chebez, 2002:68; Narosky & Yzurieta, 2010:214). Además, tiende a encaramarse en ramas secas o en árboles específicos como el ambay (Cecropia adenopus) (Narosky & Yzurieta, 2010:214).
Suelen observarse en parejas, permaneciendo posados durante períodos prolongados. Además, se señala que si una de las aves es cazada, su compañero tiende a permanecer en el mismo lugar, lo que facilita su captura, lo cual ha dado lugar a la denominación local «Juan zonzo del monte» (Pereyra, 1950:235).
Existe la posibilidad de que esta especie sea migratoria durante el verano, aunque esta información no está completamente clara. Se ha confirmado su residencia en la región de Iguazú, Misiones, lo que sugiere una distribución estable en esta área específica (Pereyra, 1950:235).
Vocalizaciones
Según lo descrito por varias fuentes (Sick, 1993:368; Rasmussen et al., 2020), su voz se caracteriza por ser muy alta y aguda para un ave de su tamaño, con secuencias descendentes de silbos melódicos, a menudo trisilábicos, que pueden ser tanto ascendentes como descendentes y que presentan notables cambios de ritmo. Se describe que estas vocalizaciones pueden sonar como «ui-ui—dibule-dibule...», y se las ha catalogado como llamadas «duras», aunque con cierto grado de melodía (Sick, 1993:368; Rasmussen et al., 2020).
ALIMENTACIÓN
En cuanto a sus hábitos alimenticios, se ha observado que frecuentan cultivos cercanos a sus dormideros, posándose en ramas débiles de árboles bajos y permaneciendo quietos durante largos períodos, a menos que detecten insectos cerca, momento en el cual los capturan y regresan a su posición anterior (Pereyra, 1950:235).
La dieta de esta especie incluye una variedad de insectos y sus larvas, así como pequeños vertebrados y materia vegetal (Rasmussen et al., 2020; Matthews & Smith, 2017:23). Específicamente, se ha observado que los insectos consumidos pertenecen a los órdenes de Orthoptera, Hymenoptera, Hemiptera, Coleoptera y Lepidoptera, con una preferencia por las larvas de Lepidoptera, Coleoptera y Himenópteros, principalmente Vespoidea. Además, se han registrado consumos de Odonata, Orthoptera, Blattodea, Auchenorrhyncha, Mantodea y, en particular de cigarras. Los adultos capturan artrópodos principalmente desde el follaje, detectando presas estacionarias tanto en el follaje como en el sustrato, y luego llevándolas de regreso a su percha favorita o directamente al nido para alimentar a los polluelos. Se ha observado que las perchas de forrajeo son utilizadas repetidamente, aunque no siempre restringidas a la proximidad del nido, ya que los adultos a veces se ausentan del área durante los periodos de observación (Matthews & Smith, 2017:23; de la Peña, 2020:29; Rasmussen et al., 2020). Bodrati & Salvador (2015:82), observaron consumo de termitas (Isoptera, Blattodea) en la provincia de Misiones.
REPRODUCCIÓN
Se reproduce durante el período de septiembre a octubre en el sur de su área de distribución (Rasmussen et al., 2020; de la Peña, 2020:29; Bodrati et al., 2010:58; Matthews & Smith, 2017:20). Excava un agujero en nidos de termitas, como se observó en un caso donde se encontró una cavidad a una altura considerable de hasta 12 metros sobre el suelo en Paraguay (Rasmussen et al., 2020).
Durante el proceso de construcción del nido, se observó la presencia de termitas dentro de la cavidad del mismo debido a la perturbación de la estructura por parte de las aves. Posteriormente, se observaron trabajos de reparación llevados a cabo por los insectos. Además, se ha documentado que las aves mantienen una cámara de anidación separada de las áreas de actividad de los insectos, posiblemente como una estrategia para evitar interferencias no deseadas en el proceso reproductivo y el cuidado de las crías (Matthews & Smith, 2017:20)
En Argentina, de la Peña, (2020:29), observó nidos en Misiones, entre noviembre y diciembre, en takurúes y en árboles emergentes; Bodrati et al. (2010:58) nidos en Parque Provincial Cruce Caballero, dpto. San Pedro Misiones, en noviembre y diciembre de 2014 y 2016, en un mismo takurú a gran altura y en un Ybyrá pytá (Peltophorum dubium). También han sido observados adultos con pichones en la provincia de Misiones (Bodrati & Cockle, 2007:291)
La puesta es de cuatro huevos de color blanco (Matthews & Smith, 2017:23; Bodrati et al., 2010:58). El período de incubación oscila entre 14 y 21 días, y el período de emplumado se extiende de 26 a 30 días aproximadamente (Matthews & Smith, 2017:20; Rasmussen et al., 2020). Además, se ha observado que tanto los adultos macho como hembra participan en la incubación, alimentación y defensa del nido, demostrando comportamientos agresivos hacia potenciales depredadores (Bodrati et al., 2010:58; de la Peña, 2020:29).
No existen datos sobre otros aspectos de la reproducción.
Depredadores
De huevos: Pteroglossus castanotis en Misiones (Cockle et al., 2016:122); Atacan a tucanes (Ramphastos dicolorus, Pteroglossus bailloni, Pteroglossus castaneus), Cyanocorax chrysops, Ictiniaplumbea, Caracara plancus, e incluso a investigadores cuando se acercan al nido (Bodrati et al., 2010:58).