Administración de Parques Nacionales. Sistema de Información de Biodiversidad. Argentina (S.I.B.) (s.f.); Ficha: Nyctidromus albicollis; recuperado de https://sib.gob.ar/especies/nyctidromus-albicollis en enero 2023.
Alvarenga, H. M. F. (1999). Sobre os hábitos de reproducto do curiango Nyctidromus albicollis (Gmelin 1798). Ararajuba, 7(1):39-40
Aragonés, J. (1997). Risk taking and flushing distance: a way of parental investment in the Pauraque (Nyctidromus albicollis). Etología, 5: 83-90.
ArgentAvis (s.f.). Ficha: Nyctidromus albicollis; recuperado de http://argentavis.com.ar/2012/sitio/especie_co.php?id=682 en enero 2023.
Baicich, P. J. & C. J. O. Harrison (1997). A Guide to the Nests, Eggs, and Nestlings of North American Birds. Second edition. Academic Press, San Diego, CA, USA. (enlace)
Barnes, R., Bartrina, L., Butchart, S. H. M., Clay, R. P., Esquivel, E. Z., Etcheverry, N. I., Lowen, J. C. & Vincent, J. (1993). Bird Surveys and Conservation in the Paraguayan Atlantic Forest. BirdLife Study Report 57. BirdLife International, Cambridge. (enlace)
Barrowclough, G. F., J. G. Groth & L. A. Mertz (2006). The RAG-1 exon in the avian order Caprimulgiformes: Phylogeny, heterozygosity, and base composition. Molecular Phylogenetics and Evolution, 41 (1):238-248. (enlace)
Belton, W. (1984). Birds of Rio Grande do Sul, Brazil. Part 1. Rheidae through Furnariidae. Bulletin of the American Museum of Natural History, 178 (4):369–636. (enlace)
Belcher, C. & G. D. Smooker (1936). Birds of the colony of Trinidad and Tobago Part III. Ibis, 66:1-35.
Bent, A. C. (1940). Life histories of North American cuckoos, goatsuckers, hummingbirds, and their allies. United States National Museum Bulletin, 176. (enlace)
Binford, L. C. (1989). A distributional survey of the birds of the Mexican state of Oaxaca. Ornithological Monographs 43. American Ornithologists’ Union, Washington, DC, USA. (enlace)
BirdLife International (2023). Species factsheet: Nyctidromus albicollis. Downloaded from http://www.birdlife.org on 30/01/2023.
Blake, E. R. (1950). Report on a collection of birds from Oaxaca, Mexico. Fieldiana, 31 (40). (enlace)
Bodrati, A. (2004). El Curiango (Nyctidromus albicollis): Presencia, Fenología y Nidificación en la Provincia de Chaco, Argentina. Revista Nuestras Aves, 47:34-36 (enlace)
Bodrati, A., Cockle, K., Segovia, J. M., Roesler, I., Areta, J. I. & Jordan, E. (2010). La avifauna del Parque Provincial Cruce Caballero, Provincia de Misiones, Argentina. Cotinga, 32:41-64. (enlace)
Bodrati, A., Areta, J. I. & White, E. (2012). La avifauna de la Posada y Reserva Puerto Bemberg, Misiones, Argentina. Nuestras Aves, 57:63-79. (enlace)
Braun, M. J. & C. J. Huddleston (2009). A molecular phylogenetic survey of caprimulgiform nightbirds illustrates the utility of non-coding sequences. Molecular Phylogenetics and Evolution, 53 (3):948-960. (enlace)
Burgos, F. G., Baldo, J. L., & Cornell, F. M. (2009). Lista de las Aves de la Provincia de Jujuy, Argentina. Secretaría de Turismo y Cultura de Jujuy. SS de Jujuy. (enlace)
Capllonch, P., Lobo, R., Ortiz, D. & R. Ovejero (2005). La avifauna de la selva de galería en el noreste de Corrientes, Argentina: Biodiversidad, patrones distribución y migración. Insugeo, Miscelánea, 14: 483-498 (enlace)
Carriker, M. A., Jr. (1910). An annotated list of the birds of Costa Rica including Cocos Island. Annals of the Carnegie Museum, 6:314-970. (enlace)
Chapman, F. M. (1929). My tropical air castle: Nature studies in Panama. New York: D. Appleton and Company. (enlace)
Chatellenaz, M. L., P. Cano, C. Saibene & H. A. Ball (2010). Inventario de las aves del Parque Nacional Mburucuyá (provincia de Corrientes, Argentina). Acta Zool. Lilloana, 54:139-160. (enlace)
Chebez, J. C. (1996). Fauna misionera, catálogo sistemático y zoogeográfico de los vertebrados de la provincia de Misiones (Argentina). Monografía No 5.; 320 pp. Ed. L.O.L.A. (Literature of Latin America). Buenos Aires. ISBN 950-9725-20-X
Chebez, J. C., N. Rey, M. Babarskas & A. G. Di Giácomo (1998). Las aves de los Parques Nacionales de Argentina. Administración de Parques Nacionales y Asociación Ornitológica del Plata. Monografía Especial L.O.L.A. Nº 12. Buenos Aires. 127 pp.
Chubb, C. (1910). The birds of Paraguay. Ibis, (9) 4:53–78. (enlace)
Cintra, R. & C. Yamashita (1990). Habitats, Abundancia e Ocorrência das Espécies de Aves do Pantanal de Poconé, Mato Grosso, Brasil. Pap. Avulsos Zool., 37 (1): 1-21. (enlace)
Cleere, N. (1998). Nightjars: A Guide to the Nightjars, Nighthawks, and Their Relatives. Yale University Press, New Haven, Connecticut, USA. (enlace)
Cockle, K. L., Bodrati, A., Lammertink, M., Bonaparte, E. B., Ferreyra, C., & Di Sallo, F. G. (2016). Predators of bird nests in the Atlantic forest of Argentina and Paraguay. The Wilson Journal of Ornithology, 128 (1):120-131. (enlace)
Coconier, E. G., López Lanús, B., Roesler, I., Moschione, F., Pearman, M., Blendinger, P., Bodrati, A., Monteleone, D., Casañas, H., Pugnali, G. & M. E. Álvarez (2007). Lista comentada de las aves silvestres de la Unidad de Gestión Acambuco. Pp: 32-103. En Coconier, E. G. (Ed.). Las aves silvestres de Acambuco, Provincia de Salta, Argentina. Temas de Nat. y Cons. No 6. Nuestras Aves A.O.P. (enlace)
Contreras, J. R. (1981). Lista preliminar de la avifauna correntina. I. No Passeriformes. Historia Natural, 2:21-28.
Contreras, J. R. (1987). Lista preliminar de la avifauna de la provincia de Formosa, República Argentina. Historia Natural, 7 (4):33-52.
Contreras, J. R., L. M. Berry, A. O. Contreras, C. C. Bertonatti & E. E. Utges (1990). Atlas Ornito-geográfico de la Provincia del Chaco – República Argentina. Volume 1. No Passeriformes. Cuadernos Técnicos “Félix de Azara” 1.
Contreras, J. R., E. R. Krauczuck, A. R. Giraudo, A. E. Johnson, A. A. Garello & Y. E. Davies (1994). Notas sobre aves de la provincia de Misiones, República Argentina. I. Nótulas Faunísticas, 53:1-13 (enlace)
Contreras, J. R., F. Agnolin, Y. E. Davies, I. Godoy, A. Giacchino & E. Ríos (2014). Atlas ornito-geográfico de la provincia de Formosa, República Argentina. (1º Ed.) Ciudad Autónoma de Buenos Aires: Fundación de Historia Natural Félix de Azara. 450 p. (enlace)
Dearborn, N. (1907). Catalogue of a Collection of Birds from Guatemala. Publications of the Field Museum of Natural History (Ornithological Series), 125: pt. 1(3). 69 pp. (enlace)
de la Peña, M. R. (1999). Aves Argentinas. Lista y Distribución. Monografía N.º 18; 244 pp. Ed. L.O.L.A. (Literature of Latin America). Buenos Aires. ISSN 0328-1620
de la Peña, M. R. (2006). Lista y Distribución de las Aves de Santa Fe y Entre Ríos. Monografía N.º 15; 137 pp. (1º Ed.). Ed. L.O.L.A. (Literature of Latin America). Buenos Aires, Argentina. ISSN 0328-1620
de la Peña, M. R. (2011). Atlas ornito-geográfico de la Provincia de Santa Fe, Argentina. Ediciones Biológica. Serie Naturaleza, Conservación y Sociedad, No 4. 500 pp.
de la Peña, M. R. (2013a). Citas, observaciones y distribución de Aves Argentinas. Serie: Naturaleza, Conservación, y Sociedad, Nº7 (1a edición). Ediciones Biológica. pp. 770.
de la Peña M. R. (2013b). Nidos y reproducción de las Aves Argentinas. Ediciones Biológica. Serie Naturaleza, Conservación y Sociedad N° 8. Santa Fe, Argentina. 590 pp. ISBN 978-987-27043-7-7
de la Peña, M. R. (2015). Aves Argentinas. Incluye nidos y huevos. 1º Ed. Santa Fe: Ediciones UNL; CABA: EUDEBA; Vol 1; 496 pp. ISBN 978-987-657-986-5
de la Peña, M. R. (2020). Aves Argentinas. Descripción, Comportamiento, Reproducción y Distribución. (Actualización). Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie) Año 2020 | N° 6 | Pág. 1-348| ISSN 0325-3856
del Hoyo, J. ed. (2020). All the Birds of the World. Lynx Edicions. Barcelona, España. 967 pp. ISBN 978-84-16728-37-4
Dickey, D. R. & A. J. van Rossem (1938). The birds of El Salvador. Field Museum of Natural History Zoological Series, Chicago, IL, USA. (enlace)
Di Giacomo, A. G. & Krapovickas, S. F. (2005). Aves de la Reserva El Bagual. Temas de Nat. y Cons. Monografía de Aves Argentinas, 4:201- 465. (enlace)
Dubs, B. (1992). Birds of Southwestern Brazil: Catalogue and Guide to the Birds of the Pantanal of Mato Grosso and its Border Areas. Betrona Verlag, Kusnacht, Switzerland. (enlace)
EcoRegistros (2023). Ficha: Nyctidromus albicollis; recuperado de https://www.ecoregistros.org/ficha/Nyctidromus-albicollis en enero 2023.
Edwards, E. P. (1983). «Nyctidromus albicollis.» In Costa Rican natural history., edited by D. H. Janzen, 590-592. Chicago, IL: Univ. of Chicago Press. (enlace)
Edwards, E. P. & R. B. Lea (1955). Birds of the Monserrate area, Chiapas, Mexico. Condor, 57:31-54. (enlace)
ffrench, R. (1991). A Guide to the Birds of Trinidad and Tobago. Revised edition. Cornell University Press, Ithaca, New York, USA. (enlace)
Foster, M. S. (1975). The overlap of molting and breeding in some tropical birds. Condor, 77:304-314. (enlace)
Foster, M. S. & N. K. Johnson (1974). Notes on birds of Costa Rica. Wilson Bulletin, 86:59-63. (enlace)
Fraga, R. M. (2001). The avifauna of Estancia San Juan Poriahú, Iberá marshes, Argentina: check-list and some natural history notes. Cotinga, 16:81-86
Friedmann, H. (1948). Birds collected by the National Geographic Society’s expeditions to northern Brazil and southern Venezuela. Proceedings of the United States National Museum, 97:373–570. (enlace)
Friedmann, H., L. Griscom & R. Moore (1950). Distributional checklist of the birds of Mexico, Part I. Pacific Coast Avifauna 29. (enlace)
Giai, A. G. (1950). Notas de viajes. El Hornero, 9 (02):121-164. (enlace)
Giraudo, A. R., J. L. Baldo & R. R. Abramson (1993). Aves observadas en el sudeste, centro y este de Misiones (República Argentina), con la mención de especies nuevas o poco conocidas para la provincia. Nótulas Faunísticas, 49:1-13. (enlace)
Giraudo, A. R. , A. Bortoluzzi & V. Arzamendia (2006). Vertebrados tetrápodos de la reserva y sitio ramsar “Esteros del Iberá” (Corrientes, Argentina): análisis de su composición y nuevos registros para especies amenazadas. Natura Neotropicalis, 37 (1-2):1-20. (enlace)
González-García, F. (1993). Avifauna de la Reserva de la Biosfera “Montes Azules”, Selva Lacandona, Chiapas, México. Acta Zool. Mexicana, 55 (1):1–86. (enlace)
Griscom, L. (1932). The distribution of bird-life in Guatemala: a contribution to a study of the origin of Central American bird-life. Bulletin of the American Museum of Natural History, New York, NY, USA. (enlace)
Hackett, S. J., R. T. Kimball, S. Reddy, R. C. K. Bowie, E. L. Braun, M. J. Braun, J. L. Chojnowski, W. A. Cox, K. L. Han, J. Harshman, C. J. Huddleston, B. D. Marks, K. J. Miglia, W. S. Moore, F. H. Sheldon, D. W. Steadman, C. C. Witt & T. Yuri (2008). A phylogenomic study of birds reveals their evolutionary history. Science, 320 (5884):1763–1768. (enlace)
Hamilton, R. D. (1948). The correct name for the “Pauraque”. Auk, 65 (1):129-130. (enlace)
Han Kin-Lan, M. B. Robbins & M. J. Braun (2010). A multi-gene estimate of phylogeny in the nightjars and nighthawks (Caprimulgidae). Molecular Phylogenetics and Evolution, 55 (2):443–453. (enlace)
Harrison, C. J. O. (1978). A Field Guide to the Nests, Eggs and Nestlings of North American Birds. Collins, Toronto, ON, Canada. 416 pp. ISBN 0-529-05484-1 (enlace)
Hartert, E. (1897). Podargidae, Caprimulgidae und Micropterygidae. Berlin: Verlag von R. Friedlander und Sohn. (enlace)
Hartman, F. A. (1961). Locomotor mechanisms in birds. Smithsonian Miscellaneous Collection, 143:1–99. (enlace)
Haverschmidt, F. & G. F. Mees (1994). Birds of Surinam. Paramaribo, Suriname: VACO N.V. (enlace)
Hayes, F. E. (1995). Status, Distribution and Biogeography of the birds of Paraguay. Monographs in Field Ornithology 1. American Birding Association, Colorado Springs, CO, USA. 230 pp. ISBN 1-878788-30-2 (enlace)
Hilty, S. L. & W. L. Brown (1986). Aves de Colombia. Princeton University Press, Princeton, NJ, USA. 1030. 294 pp. ISBN 958-33-2254-7 (enlace)
Hilty, S. L. (2002). Birds of Venezuela. Princeton: Princeton University Press. ISBN 0-691-02131-7
Howell, T. R. (1957). Birds of a second-growth rain forest area of Nicaragua. Condor, 59 (2):73–111. (enlace)
Howell, S. N. G. & S. Webb (1995). A Guide to the Birds of Mexico and Northern Central America. Oxford University Press, New York, NY, USA. 851 pp. ISBN 0-19-854013-2 (enlace)
Howell, S. N. G., C. Corben, P. Pyle & D. I. Rogers (2003). The first basic problem: a review of molt and plumage homologies. Condor, 105:635–653. (enlace)
Howell, S. N. G., C. Corben, P. Pyle & D. I. Rogers (2004). The first basic problem revisited: reply to commentaries on Howell et al. (2003). Condor, 106:206–210. (enlace)
Humphrey, P. S. & K. C. Parker (1959). An approach to the study of molts and plumages. Auk, 76 (1):1–31. (enlace)
Ingels, J. (1975). Notes on the Pauraque Nyctidromus albicollis in French Guiana. Bull. Brit. Orn. Club., 95 (3):115-116. (enlace) https://www.biodiversitylibrary.org/page/40826537
Jenkinson, M. A. & R. M. Mengel (1970). Ingestion of stones by goatsuckers (Caprimulgidae). Condor, 72:236-237. (enlace)
Kaufman, K. (1996). Lives of North American Birds. Houghton Mifflin Company, Boston & New York. (enlace)
Krauczuk, E. R. (2005). Aves do Inta-Campo Anexo ao Zaiman, e do Campus da Universidade Nacional de Misiones, Posadas, Misiones, Argentina. Atualidades Ornitológicas, 126:17-39. (enlace)
Krauczuk, E. R. (2008). Riqueza específica, abundancia y ambientes de las aves de Corpus Christi, San Ignacio, Misiones, Argentina. Lundiana: International Journal of Biodiversity, 9 (1):29-39. (enlace)
Krauczuk, E. R. (2013). Algunos aspectos de la biología de Chordeiles pusillus en Misiones, Argentina y apuntes sobre otros Caprimulgiformes. Atualidades Ornitológicas On-Line, 173:58-68 (enlace)
Kutac, E. A. & S. C. Caran (1994). Birds and Other Wildlife of South Central Texas: A Handbook. University of Texas Press, Austin, TX, USA. 203 pp. ISBN 0-292-75550-3 (enlace)
Lasiewski, R. C. & W. R. Dawson (1964). Physiological responses to temperature in the Common Nighthawk. Cóndor, 66:477-490. (enlace)
Latta, S. C. & C. A. Howell (2020). Common Pauraque (Nyctidromus albicollis), versión 1.0. In Birds of the World (A. F. Poole, Editor). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.compau.01
Lee, W. B. (1873). Ornithological Notes from the Argentine Republic. Ibis, 1873: 129-138. (enlace)
Lepage, D. (2003). Avibase-La base de datos World Bird; Ficha: Nyctidromus albicollis; recuperado de https://avibase.bsc-eoc.org/species.jsp?avibaseid=F2CB69EC613FDFE7 en enero 2023.
Lee Jones, H. & Vallely, A. C. (2001). Annotated Checklist of the Birds of Belize. Lynx Edicions. Barcelona, España. 71 pp. ISBN 84-87334-35-0
Livezey, B. C. & R. L. Zusi (2007). Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zoological Journal of the Linnean Society, 149:1–95. (enlace)
López-Lanús, B. (1997). Inventario de Aves del Parque Nacional “Río Pilcomayo” Formosa, Argentina. Monografía Especial 4. Ed. L.O.L.A. (Literature of Latin America), Buenos Aires. 78 pp. ISSN 0328-1620
López-Lanús, B., A. S. Di Giacomo, A. Azpiroz, P. Haynes, A. Galimberti, A. Keyel, A. Ocampo, R. Güller, R. Moller Jensen, M. Mattalia, H. Cardoso, C. Giarduz, G. Papini & A. G. Di Giacomo (2013). Inventario focal de fauna de las estancias La Higuera, María Concepción, La Serena y Virocay en el sitio piloto Aguapey: Corrientes, Argentina. En pp. 179-223/277-293/307-319/331-347/490,494-506: G. D. Marino, F. Miñarro, M. E. Zaccagnini y B. López-Lanús (eds.). Pastizales y sabanas del cono sur de Sudamérica: iniciativas para su conservación en la Argentina. Temas de Naturaleza y Conservación, Monografía de Aves Argentinas No 9. Aves Argentinas/AOP, Fundación Vida Silvestre Argentina e Instituto Nacional de Tecnología Agropecuaria. Buenos Aires, Argentina. ISBN: 9789874531612 (enlace)
Lowery, G. H. & Dalquest, W. W. (1951). Birds from the State of Veracruz, Mexico. University of Kansas Publications, Museum of Natural History, 3 (4). University of Kansas, Lawrence, Kansas. 119 pp. (enlace)
Manassero, M. & H. Luna (2009). Nuevos registros de aves para Santa Fe, Argentina. Parte IV. Nuestras Aves, 54:37-39. (enlace)
Marateo, G., H. Povedano & J. Alonso (2009). Inventario de las aves del Parque Nacional El Palmar, Argentina. Cotinga, 31:47-60. (enlace)
Mariaux, J. & M. J. Braun (1996). A molecular phylogenetic survey of the nightjars and allies (Caprimulgiformes) with special emphasis on the potoos (Nyctibiidae). Molecular Phylogenetics and Evolution, 6 (2):228-244. (enlace)
Mayr, G. (2010). Phylogenetic relationships of the paraphyletic ‘caprimulgiform’ birds (nightjars and allies). Journal of Zoological Systematics and Evolutionary Research, 48 (2):126-137. (enlace)
Mazar Barnett, J. & M. Pearman (2001). Lista comentada de las Aves Argentinas (1º Ed.) Lynx Edicions, Barcelona, España. 164 pp. ISBN 84-87334-32-6
Merrill, J. C. (1878). Notes on the ornithology of southern Texas, being a list of birds observed in the vicinity of Fort Brown, Texas, from February 1876 to June 1878. Proceedings of the United States National Museum, 1:118-173. (enlace)
Meyer de Schauensee, R. & W. H. Phelps (1978). A Guide to the Birds of Venezuela. Princeton University Press, Princeton, NJ, USA. (enlace)
Miller, A. H. (1963). Seasonal activity and ecology of the avifauna of an American equatorial cloud forest. University of California Publications in Zoology, 66:1–78.
Monroe, B. L., Jr. (1968). A distributional survey of the birds of Honduras. Ornithological Monographs 7. American Ornithologists’ Union, Washington, DC, USA. (enlace)
Monroy Ojeda, A., M. Grosselet, G. Ruiz & J. Nochebuenas Jaramillo (2012). Primary feather replacement on a hatch year Common Pauraque (Nyctidromus albicollis). North American Bird Bander, 37 (3):100-102. (enlace)
Moschione, F., O. Spitznagel & M. González (2013). Lista de aves de Salta. Gobierno de la Provincia de Salta. (enlace)
Narosky, T., & D. Yzurieta (2010). Birds of Argentina and Uruguay: a field guide. (16ª Ed.) Vázquez Mazzini Editores, Buenos Aires, Argentina. 432 pp. ISBN 978-987-9132-27-2
Narosky, T & J. C. Chebez (2002). Aves de Iguazú: guía para la identificación (1º Ed.). Vázquez Mazzini Editores, Buenos Aires, Argentina. 128 pp. ISBN 987-9132-04-1
Nicol, J. A. C., F. R. S. Arnott & H. J. Arnott (1974). Tapeta lucida in the eyes of goatsuckers (Caprimulgidae). Proc. R. Soc. London, Ser. B 187:349-352. (enlace)
Nores, M. & D. Yzurieta (1986). Nuevas localidades para aves argentinas. Parte 7. Hist. Nat., 6:49-52
Nores, M. (1992). Bird speciation in subtropical South America in relation to forest expansion and retraction. Auk, 109: 346-357. (enlace)
Oberholser, H. C. (1914). A monograph of the genus Chordeiles Swainson, type of a new family of goatsuckers. United States National Museum Bulletin, 86. (enlace)
Oberholser, H. C. (1974). The Bird Life of Texas. University of Texas Press, Austin, TX, USA. 1069 pp. ISBN 0-202-70711-8 (enlace)
Olrog, C. Ch. (1959). Las Aves Argentinas. Una Guía de Campo. Universidad Nacional de Tucumán. “Instituto Miguel Lillo”, Tucumán, Argentina. 343 pp.
Olrog, C. Ch. (1979). Nueva Lista de la Avifauna Argentina. Ópera Lilloana XXVII. Ministerio de Cultura y Educación. Fundación Miguel Lillo. Tucumán, Argentina. 324 pp. ISSN 0078-5245
Oniki, Y. & E. O. Willis (1982). Breeding records of birds from Manaus Brazil. Accipitridae to Caprimulgidae. Rev. Bras. Biol., 42 (4):733-740.
Parker, T. A. & J. M. Goerck (1997). The importance of national parks and biological reserves to bird conservation in the Atlantic Forest region of Brazil. In Studies in Neotropical Ornithology Honoring Ted Parker (J. V. Remsen, Editor), Ornithological Monographs, 48:527–541. (enlace)
Pautasso, A. A. (2002). Nuevos registros para aves poco citadas en Santa Fe, Argentina. Nuestras Aves, 43:19-21. (enlace)
Peters, J. L. (1940). Check-list of Birds of the World. Volume 4. Harvard University Press, Cambridge, MA, USA. (enlace)
Peterson, R. T. (1960). A Field Guide to the birds of Texas. Boston, MA: Houghton Mifflin Co. 304 pp. ISBN 0-395-08087-8 (enlace)
Planqué, R., Vellinga, W.P., Pieterse, S., Jongsma J. & de By, R. (Xeno-canto Foundation). Ficha: Nyctidromus albicollis; recuperado de https://xeno-canto.org/species/Nyctidromus-albicollis en enero 2023.
Price, J., Droege, S. & Price, A. (1995). The Summer Atlas of North American Birds. Academic Press, London. (enlace)
Pyle, P. (1995). Incomplete flight feather molt and age in certain North American non-passerines. North American Bird Bander, 20:15–26. (enlace)
Pyle, P. (1997). Identification Guide to North American Birds. Part I. Slate Creek Press, Bolinas, CA, USA.
Pyle, P. (2006). Staffelmauser and other adaptive strategies for wing molt in larger birds. Western Birds, 37:179–185. (enlace)
Quesnel, V. C. (1986). Why do nightjars sit on the road at night? Living World, J. Trin. Tob. Field Nat. Club 1985-1986:19-23. (enlace)
Quesnel, V. C. (1990). Observations on the pauraque nightjar, Nyctidromus albicollis (Gmelin), on the roads at night. Living World, J. Trin. Tob. Field Nat. Club 1989-1990:12-15. (enlace)
Reichholf, H. & J. Reichholf (1973). Mass gathering of night jars in Mato-Grosso Brazil. J. fur Ornithol., 114:503-504.
Remsen, J. V., C. D. Cadena, A. Jaramillo, M. Nores, J. F. Pacheco, J. Pérez-Emán, M. B. Robbins, F. G. Stiles, D. F. Stotz & K. J. Zimmer (2014). A classification of the bird species of South America. American Ornithologists’ Union, Louisiana State University. Available from http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm.
Ridgely, R. S. & J. Gwynne (1989). A Guide to the Birds of Panama, with Costa Rica, Nicaragua, and Honduras. 2nd Edition. Princeton University Press, Princeton, NJ, USA. 534 pp. ISBN 0-691-08529-3
Ridgway, R. (1914). The birds of North and Middle America. Bulletin of the United States National Museum, 50 part VI: 537-539 (enlace)
Rohwer, S. A. (1971). Molt and the annual cycle of the Chuck-Will’s-Widow, Caprimulgus carolinensis. Auk, 88 (3):485-519. (enlace)
Rojas, L. M., Y. Ramírez, R. McNeil, M. Mitchell & G. Marín (2004). Retinal morphology and electrophysiology of two caprimulgiformes birds: The cave-living and nocturnal Oilbird (Steatornis caripensis), and the crepuscularly and nocturnally foraging Common Pauraque (Nyctidromus albicollis). Brain, Behavior and Evolution, 64 (1):19-33. (enlace)
Root, T. (1988). Atlas of Wintering North American Birds: An Analysis of Christmas Bird Count Data. University of Chicago Press, Chicago, IL, USA. (enlace)
Rust, H. J. (1947). Migration and nesting of nighthawks in Northern Idaho. Condor, 49:177-188. (enlace)
Sandoval, L. & I. Escalante (2011). Song description and individual variation in males of the Common Pauraque (Nyctidromus albicollis). Ornitología Neotropical, 22 (2):173-185. (enlace)
Sberze, M., M. Cohn-Haft & G. Ferraz (2010). Old growth and secondary forest site occupancy by nocturnal birds in a neotropical landscape. Animal Conservation, 13 (1):3-11. (enlace)
Scholander, P. F., R. Hock, V. Walters & L. Irving (1950). Adaptation to cold in arctic and tropical mammals and birds in relation to body temperature, insulation, and basal metabolic rate. Biological Bulletin, 99:259-271. (enlace)
Schulenberg, T. S., Stotz, D. F. Lane, D. F., O’Neill, J. P., Parker III, T. A. & Brack Egg, A. (2010). Birds of Perú.Princeton University Press. 664 pp. ISBN: 978-0-691-13023-1
Sibley, C. G. & J. E. Ahlquist (1990). Phylogeny and Classification of Birds: A Study in Molecular Evolution. Yale University Press, New Haven, CT, USA.
Sick, H. (1997). Ornitologia Brasileira. Editora Nova Fronteira, Rio de Janeiro, Brazil. 912 pp. ISBN 85-209-0816-0
Sigurdsson, S. & J. Cracraft (2014). Deciphering the diversity and history of New World nightjars (Aves: Caprimulgidae) using molecular phylogenetics. Zoological Journal of the Linnean Society, 170:506-545. (enlace)
Skoruppa, M. K., M. C. Woodin & G. Blacklock (2009). Species richness, relative abundance, and habitat associations of nocturnal birds along the Rio Grande in southern Texas. Southwestern Naturalist, 54 (3):317-323. (enlace)
Skutch, A. F. (1972). Studies of tropical American birds. Publications of the Nuttall Ornithological Club 10. (enlace)
Skutch, A. F. (1976). Parent birds and their young. Austin: Univ. of Texas Press. 503 pp. ISBN 0-292-76424-3 (enlace)
Skutch, A. F. (1983). Birds of Tropical America. University of Texas Press, Austin, TX, USA. 305 PP. ISBN 0-292-74634-2 (enlace)
Slud, P. (1960). The birds of finca ‘La Selva,’ Costa Rica: a tropical wet forest locality. Bulletin of the American Museum of Natural History, 121:49–148. (enlace)
Slud, P. (1964). The birds of Costa Rica: distribution and ecology. Bulletin of the American Museum of Natural History, 128:1–430. (enlace)
Slud, P. (1964) The birds of Costa Rica: distribution and ecology. Bulletin of the American Museum of Natural History, 128. (enlace)
Slud, P. (1980). The birds of Hacienda Palo Verde, Guanacaste, Costa Rica. Smithson. Contrib. Zool., 292. (enlace)
Smithe, F. B. (1969). Las aves de Tikal. Guatemala: Litografia Byron Zadik y cia.
Snyder, D. E. (1966). The birds of Guyana (formerly British Guyana). Salem, MA: Peabody Mus. (enlace)
Stiles, F. G. & A. F. Skutch (1989). A Guide to the Birds of Costa Rica. Cornell University Press, Ithaca, NY, USA. 511 pp. ISBN 0-8014-2287-6 (enlace)
Stotz, D. F., J. W. Fitzpatrick, T. A. Parker & D. K. Moskovits (1996). Neotropical Birds: Ecology and Conservation. University of Chicago Press, Chicago, IL, USA, and London, UK. 478 pp. ISBN 0-226-77630-1
Terrill, S. B. & L. S. Terrill (1986). Common pauraque (Nyctidromus albicollis) record from Sonora, Mexico. American Birds, 40 (3):430. (enlace)
Thurber, W. A. (2003). Behavioral notes on the Common Pauraque (Nyctidromus albicollis). Orn. Neotrop, 14:99-105. (enlace)
Todd, W. E. C. & M. A. Carriker (1922) The birds of the Santa Marta region of Colombia: a study in altitudinal distribution. Annals of the Carnegie Museum, 14. (enlace)
Tweit, R. C. (2014). Common Pauraque. Texas A&M University (enlace)
Van Rossem, A. J. (1927). Eye shine in birds, with notes on the feeding habits of some goatsuckers. Condor, 29:2528. (enlace)
Vasconcelos, M. F., Figueredo, C. C., Carvalho, H. Á. & Neto, S. D. A. (2003). Observações sobre a reprodução do curiango, Nyctidromus albicollis (Gmelin, 1789),(Aves: Caprimulgiformes) no estado de Minas Gerais, Brasil. Lundiana: International Journal of Biodiversity, 4 (2):141-147. (enlace)
Vellinga, W. P., J. N. M. Flanagan & T. R. Mark (2004). New and interesting records of birds from Ayabaca province, Piura, north-west Peru. Bulletin of the British Ornithologists’ Club, 124 (2):124-142. (enlace)
West, S. (1981). Range extension of the pauraque in Texas. Bulletin of the Texas Ornithological Society, 14:25-26. (enlace)
Wetmore, A. (1944). A collection of birds from northern Guanacaste, Costa Rica. Proceedings of the United States National Museum, 95:25-80. (enlace)
Wetmore, A. (1968). The birds of the Republic of Panama, Part 2. Columbidae (Pigeons) to the Picidae (Woodpeckers). Smithsonian Miscellaneous Collections, 150 (2):1–605. (enlace)
Zimmer, J. T. (1930). Birds of the Marshall Field Peruvian Expedition, 1922–1923. Field Museum of Natural History Zoological Series, 17:233–480. (enlace)
ESTADO DE CONSERVACIÓN SEGÚN BIRDLIFE INTERNACIONAL: PREOCUPACIÓN MENOR
Esta especie tiene un rango de distribución extremadamente grande y por lo tanto no se aproxima a los umbrales de vulnerabilidad bajo criterio de tamaño de la extensión de la población (Extensión < 20.000 km² combinado con una declinación o fluctuación del tamaño de la extensión, extensión/calidad del hábitat, o tamaño de la población y un pequeño número de localizaciones o con fragmentación grave). La tendencia de la población parece ser estable, y por lo tanto la especie no alcanza los umbrales de Vulnerabilidad bajo el criterio de tendencia poblacional (disminución > 30% en diez años o en tres generaciones). El tamaño de la población no ha sido cuantificada, pero no se cree que alcance los umbrales de Vulnerabilidad bajo el criterio de tamaño de la población (<10.000 individuos maduros con una disminución continua estimada > del 10% en diez años o en tres generaciones o con una estructura de población específica). Por estas razones, la especie es evaluada como “común” y de “preocupación menor” (Stotz et al., 1996:172).
Se estima que la población global es estable y de cerca de 20 millones de individuos maduros (S.I.B).
Efectos de la actividad humana
Cazados por los nativos locales en mato Grosso, Brasil. Existen varias reseñas de atropellos por vehículos en los caminos, mientras permanecen posados (Reichholf & Reichholf, 1973:503), y abandono de nidos tras perturbaciones por los humanos en Brasil (Vasconcelos et al.,2003:144). Muchos huevos y ejemplares jóvenes destruidos por el fuego asociado a la agricultura de tala y quema (Skutch, 1972:19).
Estado de Conservación en Argentina: (S.I.B.)
No Amenazada (NA): según AA y SAyDS, 2008 (Aves Argentinas y Secretaría de Ambiente y Desarrollo Sustentable ). López-Lanús, B., P. Grilli, E. Coconier, A. Di Giacomo y R. Banchs. Categorización de las aves de la Argentina según su estado de conservación. Informe de Aves Argentinas /AOP y Secretaría de Ambiente y Desarrollo Sustentable. Buenos Aires, Argentina. 1 ed. AOP: Buenos Aires, Argentina No Amenazada (NA): según MADS y AA, 2017 (Ministerio de Ambiente y Desarrollo Sustentable de la Nación y Aves Argentinas ). Categorización de las Aves de la Argentina según su estado de conservación. Informe del Ministerio de Ambiente y Desarrollo Sustentable de la Nación y de Aves Argentinas. 146 p. 2017 No Amenazada (NA): según SAyDS, 2010 (Secretaría de Ambiente y Desarrollo Sustentable ). Categorización de aves autóctonas de Argentina. Lista de aves no registradas, endémicas y exóticas de Argentina (Res 348). 2010 Preocupación Menor (LC): según UICN (API) (Unión Internacional para la Conservación de la Naturaleza ). Lista Roja de la UICN publicada en https://www.iucnredlist.org. Actualización en tiempo real
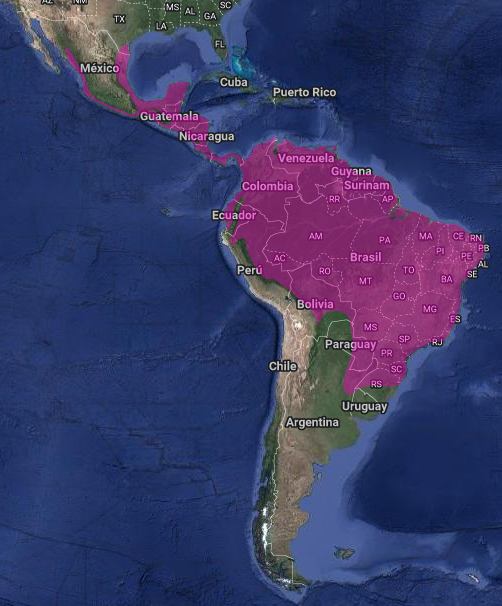
InfoNatura: Animales y Ecosistemas de América Latina [aplicación web]. 2007. Versión 5.0. Arlington, Virginia (Estados Unidos): NatureServe. Disponible: http://www.natureserve.org/infonatura.Se reconocen seis subespecies (de la Peña, 2020:229) con un amplio rango de distribución (Stotz et al., 1996:316). Abarcan desde el extremo sur del Estado de Texas (por encima de los 195 msnm) (Price & Price, 1995:280; Root, 1988:129; Oberholser, 1974:478; Kaufman, 1996:325) y valle del Río Grande, México por la pendiente del Caribe (Bent, 1940:199) desde Tamaulipas y este de Nuevo León (Lowery & Dalquest, 1951:579), y hacia el sur a lo largo de la pendiente del Pacífico (Terrill & Terrill, 1986:430; Binford, 1989:135; Blake, 1950:401; González-García, 1993:38; Howell & Webb, 1995:375). La distribución continúa hacia el sur a través de América Central en, Belice, Guatemala (por encima de los 1,740 msnm; Griscom, 1932:190; Skutch, 1972:3; Dearborn, 1907:95; Land, 1962:273), pero ausente en las tierras altas del sur de Guatemala y la adyacente Chiapas (Howell & Webb, 1995:375), Honduras (por encima de los 1.800 msnm, más común en la pendiente del Caribe (Monroe, 1968:359), El Salvador (por encima de los 1.100 msnm) (Van Rossem, 1927:27; Dickey & van Rossem 1938:242; Thurber, 2003:100), Nicaragua, Costa Rica (por encima de los 2.700 msnm) (Slud, 1980:20; Stiles & Skutch, 1989:200; Edwards, 1983:590; Carriker, 1910:502 por encima de los 2.700 msnm), y Panamá (Wetmore, 1968:208; Chapman, 1929:22) (por encima de los 1.800 msnm y localmente en la pendiente del Caribe (Ridgely & Gwynne, 1989:194), También en Colombia (donde se lo ha registrado a 2.300 msnm) (Todd & Carriker, 1922:218; Hilty & Brown, 1986:294), Venezuela (por encima de los 1.400 msnm) (Friedmann, 1948:414; Hilty, 2002:376), Trinidad y Tobago (Trinidad y Tobago (ffrench, 1991:208), Surinam (Haverschmidt & Mees, 1994:166; Lee Jones & Vallely, 2001:26) y sur de Guayana francesa (Ingels, 1975:115), Guayana (Snyder, 1966:126), al oeste y NO de los Andes en Perú (Zimmer, 1930:268; Vellinga et al., 2004:133), al este de los Andes a través de Ecuador, este de Perú (Schulenberg et al., 2010:202), norte y este de Bolivia, todo Brasil (Olrog, 1979:134; Belton, 1984:561; Dubs, 1992:53; Sick, 1997:418), y Paraguay (Hayes, 1995:66 y 144; Barnes et al., 1993:138 y 143; Chubb, 1910:267) hasta el norte de Argentina (Mazar Barnett & Pearman, 2001:48) donde se ha registrado en las provincias de Salta, Jujuy, Corrientes, Formosa, Chaco, Misiones, Entre Ríos y noreste de Santa Fe (Olrog, 1959:155; Olrog, 1979:134; de la Peña, 1999:90; de la Peña, 2013b:195; de la Peña, 2015:328) y Uruguay.
En Argentina (Contreras et al., 2014:310) la raza N. a. derbyanus (la más sureña de las seis subespecies) está presente en el extremo norte de Salta (Olrog, 1979:134; de la Peña, 1999:90), este de Formosa,Jujuy (Nores e Yzurieta, 1986; Contreras et al., 1990; Di Giácomo & Krapovickas, 2005:300) y de Chaco (Chebez et al., 1998; Bodrati, 2004), norte de Corrientes (de la Peña, 2013a:287), noroeste de Santa Fe (Giai, 1950:163; Narosky & Yzurieta, 2010:194; de la Peña, 2013a:287; de la Peña, 2006:64; de la Peña, 2020:229), Entre Ríos donde hay un registro accidental muy antiguo, observado en bancos de Arroyo el Gato 7-8 leguas (35- 40 km) al NO de Gualeguaychú de julio a febrero (Lee, 1873:134; Olrog, 1979:134; de la Peña, 2015:329) y Misiones (Giai, 1950:164). (Citas y coordenadas en los enlaces)
HÁBITAT
Frecuenta estratos bajos de 0-1700 msnm, (ocasionalmente hasta los 2.300 msnm) en la pendiente árida del Pacífico, Golfo del Caribe, tierras bajas del Chocó, Costa ecuatorial del Pacífico, Norte y sur de Amazonia, América Central y norte de América del Sur (Stotz et al., 1996:172). Escaso en la mayoría de las zonas forestadas, no frecuenta bosques espesos (Stiles & Skutch, 1989:201). Parece preferir ambientes antrópicos como chacras, caminos, bosques secundarios y abiertos (Di Giácomo & Krapovickas, 2005:300). Por lo general se encuentra cerca de bosques o selvas en galería en óptimo estado de conservación (Bodrati, 2004:35) Bosques tropicales y subtropicales (Oberholser, 1974:478; Contreras et al., 2014:309), montes, zona de matorrales y de cultivo (café en Costa Rica) (Stiles & Skutch, 1989:201) (palma en El Salvador (Dickey & van Rossem, 1938:186), llanos, pantanos, manglares y bosque de segundo crecimiento en Venezuela (Meyer de Schauensee & Phelps 1978:124), sabanas y bordes de bosques en Trinidad (ffrench, 1991:208), capoeiras (Sick, 1997:418) y bosque secundario al norte de Manaos en Brasil (Sberze et al., 2010:8). En Texas, en el Valle del Río Grande, se lo encuentra frecuentemente cerca del agua (Skoruppa et al., 2009:319). Ecorregiones: Selva Paranaense, Selva de Yungas y Chaco (Mazar Barnett & Pearman, 2001:48).
De hábitos crepusculares y nocturnos, excepto cuando se ve perturbado durante el día (Contreras et al., 2014:309; de la Peña, 2020:228). Los individuos se desplazan de forma aislada (ffrench, 1991:209) o en pequeños grupos familiares (Skutch, 1972:20; Quesnel, 1990:13), si bien es común que se produzcan aglomeraciones mayores durante la búsqueda de alimento (Reichholf & Reichholf, 1973; citados por Latta & Howell, 2015)
Se ha observado que, tanto las llamadas como la actividad nocturna, aumentan en la primavera a medida que la luna crece, y durante las noches iluminadas por la luna (Quesnel, 1990:14; Thurber, 2003:102).
Durante el día, los individuos descansan en el suelo entre la maleza y la cubierta del suelo; su plumaje crÍptico hace que su detección sea extremadamente difícil (Contreras et al., 2014:309). Cuando descansan, tienen la cabeza encogida y el cuerpo tendido, como otros caprimúlgidos, pero cuando se les acerca, pueden extender la cabeza y luego retraerla o levantar el cuerpo y luego volver a caer. Cuando se les molesta, los pájaros parecen muy alertas y se espantan fácilmente o, más raramente, caminan unos pasos. Ocasionalmente, después de ser espantados, se posan en un tronco o una rama baja, donde descansan en posición longitudinal para ayudar a ocultarse (Wetmore, 1968:210). No se conocen cambios en los patrones de actividad con la reproducción. A menudo está inactivo en climas fríos y por lo tanto se cree que está ausente (Peterson, 1960:135). Los sitios de descanso incluyen plantaciones, claros arbustivos, matorrales, bordes de bosques sobrecrecidos y aberturas hechas por humanos en regiones densamente boscosas (Slud, 1964:137). En la temporada de lluvias y cuando las áreas bajas están inundadas, generalmente seleccionan lugares de descanso ligeramente elevados a 3-10 m sobre el suelo (Wetmore, 1968:210). En la temporada de no reproducción, muchos individuos pueden instalarse en el mismo matorral o parche de maleza.
Con patas cortas pero bien desarrolladas, y más largas que las de sus parientes inmediatos, salta verticalmente 0.5 m del suelo (o ocasionalmente desde una percha en una roca o tronco) para atrapar insectos voladores bajos; también puede mover las alas durante el salto (Dickey & van Rossem, 1938:243). Ocasionalmente corre en el suelo para forrajear (Hartert, 1897:32) o puede caminar una corta distancia (Edwards, 1983:590). Mientras canta, puede levantarse «ligeramente en sus pies» sin perder contacto con la superficie de apoyo (Skutch, 1972:4). También se observó que flexionaba las piernas repetidamente en un movimiento de balanceo mientras estaba en el suelo (Hartman, 1961:11).
Durante el vuelo generalmente se mantiene a una distancia de 3 m del suelo (Oberholser 1974:478). Vuela al atardecer desde los sitios de nidificación diurnos hasta los terrenos de alimentación. Ocasionalmente realiza picadas circulares, para atrapar insectos por la noche, desde terrenos abiertos, rocas o postes de alambrado. Estos vuelos cortos generalmente terminan en el punto de partida o cerca de él (Skutch, 1972:4). Cuando son asustados, los individuos se alejan de forma errática y silenciosa (Oberholser 1974:478, Ridgely & Gwynne, 1989:194). El vuelo continúa a través o debajo de árboles y arbustos, no sobre ellos, hasta que el ave se posa en el suelo, a menudo cerca de su posición original (Skutch, 1972:4), a 15-20 m de distancia (Edwards, 1983:590-591), u ocasionalmente en una percha baja (Stiles & Skutch, 1989:201).
Los adultos, cuando incuban, es raro que se acicalen mientras están en el nido (quizás para mantener la discreción). Un individuo empapado por la lluvia salió del nido para acicalarse y rascarse cuando salió el sol (Skutch, 1972:13). El pie tiene los dedos altamente modificados que pueden ser usados para peinar las vibrisas (Edwards, 1983:590). Bostezan durante la incubación exponiendo el interior de la boca (Skutch, 1972:13).
VOCALIZACIONES
Las vocalizaciones de esta especie suelen ser emitidas casi exclusivamente durante la noche, especialmente al atardecer y al amanecer (Quesnel, 1990:14); raramente se escuchan llamadas durante el día (Wetmore, 1968:210). La vocalización aumenta en las noches claras de luna llena (Oberholser 1974:478; Quesnel, 1990:14); sin embargo, se reduce con la ausencia de luna, el frío, las nubes, la lluvia y el viento (Oberholser 1974:478). El canto, parecido a un silbido, está formado por 4 sílabas emitidas rápidamente en modulación ascendente de frecuencia y luego marcadamente descendente hacia el final que repiten esporádicamente (de la Peña, 2020:228)
Se presume que se producen desde el suelo o desde una percha baja. Pueden usar senderos para emitir sus cantos o usar otras perchas favoritas (Dickey & van Rossem, 1938:243).
Varios tipos de vocalizaciones. El llamado más distintivo de la especie, generalmente se describe como un grito extraño y afligido, o un silbido áspero o ronco, traducido como «¿Who are you?»….“Whaup, whaup-hoo-yoo”, generalmente traducido como «¿Quién eres?» (Snyder, 1966:126, ffrench, 1991:209) o «hip-hip hip-hip hip-hip-hooray», pero existe una variación sustancial en las descripciones. No se sabe si la variación refleja diferencias geográficas en la canción o en las interpretaciones humanas.
Los diferentes autores lo describen así: en Texas, como un silbido áspero y brumoso, «pur pur perp pur-wheeeeeer», que se acelera a medida que avanza la noche (Oberholser 1974:478); en Costa Rica, un ronco o tembloroso «who-whick who-whick whick wick-wick-wik wik wip WHEEEEUR» o «kw-kw, kw-kw, kw-kw. kw, kw, kwah-REE-O», o un repetido, incesante y ronco «krrweeo» o «cuyeer» (Stiles & Skutch, 1989:201); en Venezuela, un áspero y fuertemente acentuado «whoo-oo-WE-oo» y un repetido «whip-whip-whip-whip» (Meyer de Schauensee & Phelps, 1978:125; Cleere, 1998:184); en Trinidad, un similar, agudo «hip-hip-hip» (ffrench, 1991:209); en Surinam, un silbido sibilante, «wook, wook, wook, wook, wook-a-whee-ooh, a-wee-ooh, a-wee-ooh, a-wee-ooh» (Haverschmidt & Mees 1994:166); y en Brasil, la canción se describe como «go-bee-oo, gril-woo, cohrianGOO» (Sick, 1997:418). En Costa Rica se estudiaron canciones de 17 machos individuales y los autores demostraron que variaban significativamente en frecuencia y otras características; algunos machos cantaban solos, otros como parte de grupos (Sandoval & Escalante, 2011:182). Aunque intentaron controlar todas las posibles causas externas de variación en las características de la canción durante el diseño de la muestra y las sesiones de grabación, no llegaron a conclusiones definitivas porque no compararon la canción del mismo individuo entre múltiples sesiones de grabación, como se propone en el nuevo enfoque para demostrar la singularidad individual. La variación masculina que encontraron podría usarse como una señal para permitir el reconocimiento de machos individuales porlas hembras o por otros machos. Encontraron que los machos en grupos tienden a cantar canciones con características más similares que con grupos distantes. Se necesitan más estudios con machos marcados para determinar si las hembras prefieren características de canción masculina particulares, y si las preferencias femeninas podrían explicar las diferencias estructurales y los patrones de canción de los machos de esta especie. También es importante determinar el papel de la canción en las interacciones macho-macho, cómo los machos forman los grupos y si estos grupos son estables a través del tiempo.
De acuerdo con Thurber (2003:101), existe una aparente imitación de canciones entre vecinos en El Salvador. Esta afirmación necesita ser confirmada en otros lugares.
Emiten diferentes «llamadas», a menudo descritas como gruñidos, notas guturales y gargarismos similares al croar de una rana (Slud, 1964:137), pero también un «waa-oo» nasal de tono más bajo y «ba-bacoorau» en Brasil. Asimismo, se registra un «whoo» soprano de tono alto y claro, además de un «llamado muy diferente», descrito como «muy bajo» y que seguramente se pierde a distancias mayores a 3 metros (Snyder, 1966:126; Stiles & Skutch, 1989:201; ffrench, 1991:209; Sick, 1997:418; Skutch, 1972:16; Latta & Howell, 2015). No se conoce la función de estos sonidos.
Emplean un llamado bajo y cacareante para atraer a los polluelos (Bent, 1940:201; Cleere, 1998:184).
Durante la época de apareamiento, la mayoría de las vocalizaciones están relacionadas con los eventos reproductivos (Wetmore, 1968:210; Quesnel, 1990:12; Cleere, 1998:184 ). Tienden a disminuir sus cantos durante la muda y la temporada no reproductiva (Oberholser, 1974:478; Thurber, 2003:102; Bodrati, 2004:34), especialmente entre julio y septiembre (Quesnel, 1990:12) o julio y octubre (Skutch, 1972:6). En América Central, parecen ser particularmente sensibles a la temporada de lluvias, cuya duración varía según la región, con poco canto incluso en las noches más luminosas de luna llena (Skutch, 1972:6).
TERRITORIALIDAD
No se observó comportamiento territorial interespecífico. El uso de caminos puede estar asociado con el establecimiento de territorios (Quesnel 1990:12), pero la falta de predominio de los machos sugiere otros usos como el de alimentación (Quesnel, 1990:14).
ALIMENTACIÓN
Se alimenta principalmente de coleópteros, comúnmente conocidos como escarabajos. Principalmente de la familia Scarabaeidae, pero también otros como cascarudos (Elateridae), escarabajos tigre (Cicindelinae), escarabajos barrenadores de la corteza (Ostomidae), escarabajos de cuernos largos (Cerambycidae), luciérnagas (Lampyridae), barrenadores de ramas (Bostrichidae).
Insectos como la hormiga león (Myrmeleontidae), chinches (Pentatomidae), langostas (Acrididae), mariposas y polillas (Lepidoptera) y abejas y avispas (Hymenoptera) (Bent, 1940:203). En Panamá, Surinam y Costa Rica, también se alimenta coleópteros como «bessbugs» (Passalidae), escarabajos picudos o gorgojos (Curculionidae), e insectos como moscas (Diptera), mantis religiosa (Mantispidae), saltamontes (Acrididae), zancudos (Tipulidae) y escarabajos del barro (Heteroceridae) (Wetmore, 1968:210; Haverschmidt & Mees, 1994:166; Edwards, 1983:591). En Yucatán, un joven Pauraque fue encontrado ingiriendo piedras (Jenkinson & Mengel, 1970:237), mientras que en Panamá se hallaron cabezas duras e indigeribles de escarabajos y «pequeñas semillas blancas duras» en dos estómagos. Estas semillas y piedras pueden servir para ayudar a triturar los alimentos.
Generalmente se alimenta en claros y áreas abiertas al anochecer y durante la noche, incluyendo carreteras (Van Rossem, 1927:27; Quesnel, 1990:12), sabanas (ffrench, 1991:208), campos de maíz (Van Rossem, 1927:27), bordes de arroyos, y ocasionalmente en huecos en áreas boscosas (Wetmore, 1968:210). Puede haber una variación estacional en los lugares de alimentación en algunas regiones (Foster & Johnson, 1974), ya que en Costa Rica los únicos pájaros observados en julio y agosto se encontraban en el bosque ribereño del río. Reicholf & Reicholf (1973, citados por Latta & Howell, 2015) informaron sobre un gran grupo de aves alimentándose (542 individuos) en Mato Grosso, Brasil, que incluía 5 especies de caprimúlgidos: 20 Hydropsalis brasiliana, 10 Caprimulgus parvulus, 2 C. rufus, 4 Podager nacunda y 2 Nyctibius griseus. Todas estas especies fueron encontradas juntas al anochecer a lo largo de 100 km de un camino de tierra.
Atrapa sus presas y se alimenta principalmente de forma terrestre; vuela muy poco mientras recolecta alimento. Aparece brincando, saltando o, en contadas ocasiones, corriendo por el suelo (Oberholser, 1974:478; Van Rossem, 1927:27; Hartert, 1897:32). Puede lograr saltar más de 0,5 m, con ayuda de un ligero giro de sus alas (Dickey & van Rossem, 1938:243). También puede brincar (Slud, 1964:137) o «sacudirse» (Meyer de Schauensee & Phelps, 1978:124) cerca del suelo para capturar insectos voladores de baja altura, o hacer vuelos cortos, aunque generalmente se limita a recorrer entre 0,3 y 3,0 m en el aire (Oberholser, 1974:478). Cuando forrajea en el aire, generalmente realiza vuelos bajos, breves y circulares para atrapar insectos voladores desde el suelo o desde una percha, roca, tronco, rama o poste cercano. Durante la temporada de lluvias en Panamá, cuando las zonas bajas están inundadas, es más probable que se posen fuera del suelo (3-10 m) (Wetmore, 1968:210). En Trinidad, comienza a cazar aproximadamente 20-25 minutos después del atardecer y parece ser más activo con la luna llena (Quesnel, 1990:14). Sin embargo, en Guatemala se ha observado que es más activo durante los periodos de luna menguante que en los de luna llena o en la oscuridad (Smithe, 1969). Se sabe que también caza bajo luces artificiales (Thurber, 2003:103).
Esta especie se caracteriza por tener una boca con una abertura muy grande, lo que facilita el forrajeo. Se presume que usa la vista para detectar insectos por silueteado o iluminación trasera de la presa contra el cielo. La presencia de un tapetum lucidum (Nicol et al., 1974:351), una estructura reflectante dentro de la coroides del ojo que mejora la visión nocturna al reflejar la luz hacia la retina, sugiere que los ojos están adaptados a condiciones de poca luz. La estructura retiniana del ojo (altamente dominada por bastones) ha sido bien estudiada por Rojas et al. (2004:19), y muestra una sensibilidad retiniana alta. No existen evidencias de que esta especie utilice la ecolocalización para la captura de alimentos.
METABOLISMO
Según Scholander et al. (1950:264) y Lasiewski & Dawson (1964:484), los Pauraques, como algunos otros Caprimúlgidos, tienen tasas metabólicas básicas más bajas que la mayoría de las otras aves de un tamaño comparable. La tasa metabólica basal del Pauraque se ha informado como 1.5-2.3 cc 02/g/h (Scholander et al. 1950:267).
En respuesta al estrés térmico, muchos Caprimúlgidos, incluyendo el Pauraque, aumentan la disipación de calor al jadear (reforzado por la gran apertura de la boca) y con el temblor gular (Skutch, 1972:12).
REPRODUCCIÓN
En Texas, se reproduce desde principios de marzo hasta julio-agosto (Bent, 1940:205; Cleere, 1998:185; Oberholser, 1974:478); en Trinidad y Tobago entre febrero y julio (Belcher & Smooker, 1936, citados por Latta & Howell, 2015; Quesnel, 1990:12; Cleere, 1998:185); entre febrero y mayo/junio en Panamá (Wetmore, 1968:211; french, 1991:209; Cleere, 1998:185); entre enero y mayo en Colombia (Hilty & Brown, 1986:294; Cleere, 1998:185); a principios de abril-mayo en El Salvador (Dickey & van Rossem, 1938:243); entre febrero y abril/mayo en Costa Rica (Stiles & Skutch, 1989:201; Cleere, 1998:185). Huevos encontrados en Surinam en todos los meses excepto febrero; el pico es mayo-septiembre (Haverschmidt & Mees, 1994:166; Cleere, 1998:185); agosto a noviembre en el sureste de Brasil (Vasconcelos et al., 2003:145) y según Cleere, 1998:185 septiembre?-enero?. No se ha registrado una segunda puesta (Harrison, 1978:181).
Esencialmente, no hay estructura de nido, aunque ocasionalmente se puede formar una depresión parecida a un platillo sobre el sustrato; los huevos se depositan simplemente en el suelo, en terrenos desnudos y planos, pero pueden anidar sobre hojas caídas (ffrench, 1991:209; Haverschmidt & Mees, 1994:166; Baicich & Harrison, 1997:193; Vasconcelos et al., 2003:142). Se han observado nidos tanto en la sombra como en el sol directo (Dickey & van Rossem, 1938:243-244; Stiles & Skutch, 1989:201).
Los nidos se encuentran dispersos entre los matorrales, bosques de tierras bajas, pequeños claros en las plantaciones, bordes de campos o a lo largo de los bordes más lejanos de los claros o senderos. No anidan en los matorrales espesos, como los que utilizan como refugio durante el invierno, sino que buscan terrenos más altos y abiertos evitando las colinas rocosas (Bent, 1940:200). En hábitats más abiertos, el nido puede estar oculto por un arbusto, generalmente entre arbustos dispersos y cactus. En El Salvador, los nidos se encuentran tanto en sombra como en sol directo (Dickey & van Rossem, 1938:243-244); en Costa Rica, el nido suele estar en un lugar sombreado de un sitio bastante abierto (Stiles & Skutch, 1989:201).
De acuerdo con Oberholser (1974:478), Howell y Webb (1995:375), Edwards (1983:592), Smithe (1969, citado por Latta y Howell, 2015), ffrench (1991:209), De la Peña (1990) y Hilty y Brown (1986:294), se han registrado puestas, generalmente de dos huevos en, Texas, México, Costa Rica, Guatemala, Trinidad/Tobago, Argentina y Colombia. Por otra parte, Haverschmidt y Mees (1994:166) reportan que, en Suriname, de doce puestas, solo una fue de dos huevos, mientras que las once restantes consistieron en uno solo.
Huevos elípticos o sub-elípticos (Wetmore, 1968:211; Harrison, 1978:181) de superficie lisa, sin brillo o ligeramente brillante. Color crema, salmón o más brillante que el tono vináceo, con marcas dispersas o densas de color marrón rojizo, canela y/o rosa, gris, y lila pálido; ocasionalmente con algunas líneas negras o con marcas concentradas alrededor del extremo más grande. Los detalles se encuentran en Vasconcelos et al. 2003.
El intervalo entre la puesta de los huevos fue de al menos 2 días (Skutch, 1972:8; Skutch, 1976:137). En cuanto a la incubación la información es escasa: en Costa Rica, algunos adultos fueron observados sentados sobre el primer huevo antes de la puesta del segundo (Skutch, 1972:8).
Se estima que el tiempo de incubación es de 10-20 días.
Los huevos se mantienen casi constantemente cubiertos, tal vez para reducir la pérdida de color que los haría visibles a los depredadores (Skutch, 1976:146). Macho y hembra alternan la incubación durante el día, generalmente pasando 2-3 horas en cada turno, pero se han registrado turnos de incubación de 6 horas (Skutch, 1972:9; Skutch, 1976:174; Edwards, 1983:592; Vasconcelos et al., 2003:143). El macho generalmente incuba durante el día, aunque puede descuidar el nido durante horas por la tarde. La incubación nocturna es difícil de observar, pero es realizada principalmente por la hembra (Skutch, 1972:11).
Posicionan los huevos con el pico para luego, mediante movimientos laterales del cuerpo, colocarse sobre ellos. Estos movimientos laterales continúan a intervalos ampliamente espaciados, pero pueden aumentar después de la lluvia o bajo el sol directo. Durante la lluvia, algunos individuos se sientan más alto en el nido. Los párpados del adulto están en constante movimiento, cambiando el diámetro de la pupila mientras incuba (Skutch, 1976:175). Los individuos que incuban también pueden bostezar, abrir y cerrar el pico, hacer temblar la gular o producir sonidos de «clic» (Skutch, 1972:13).
Puede mudar de sitio los huevos si es molestado (Euler, en Alvarenga, 1999:39); sin embargo Skutch (1972:21 y 1976:148) observó que los padres nunca movieron los huevos desde su ubicación original, en respuesta a los múltiples disturbios ocasionados (por ejemplo, visitar el nido, medir los huevos, erigir pantallas cerca de los nidos).
Crías semi-precociales (Skutch, 1976:230; Baicich & Harrison, 1997:193) cubiertos casi completamente por una larga y suave capa de plumón salmón oscuro con puntas negras (Skutch, 1972:15; Bodrati, 2004:35; de la Peña, 2020:229).
No hay datos de crecimiento. Los ojos de los polluelos se abren parcialmente después de la eclosión. Los pichones pueden moverse a corta distancia, incluso el mismo día de haber nacido (Cleere, 1998:185). Cambian de sitio en respuesta a las llamadas de los adultos (Sick, 1997:416; Baicich & Harrison, 1997:193; Cleere, 1998:186). Los polluelos pían a los 2-3 días (Skutch, 1972:16; Cleere, 1998:184).
Los jóvenes son alimentados por la noche, presumiblemente por ambos padres (ffrench, 1991:209). La punta estrecha del pico se inserta en la boca del polluelo, donde los insectos son regurgitados (Harrison, 1978:181). Los jóvenes son capaces de estirar sus cuerpos y cuellos para recibir comida (Skutch, 1972:16).
Son cuidados por ambos adultos. Los polluelos permanecen con sus padres hasta algún tiempo después de que estén completamente crecidos y sean capaces de volar (Skutch, 1972:21). Los jóvenes pueden acompañar a sus padres a áreas de alimentación, como lo ha registrado Quesnel (Quesnel, 1990:12) que observó grupos de 3 aves alimentándose juntas, en pleno verano, en varias ocasiones. Se han observado grupos familiares de 3-4 individuos, a finales de agosto, en El Salvador (Dickey & van Rossem, 1938:244) y Costa Rica (Foster & Johnson, 1974, citados por Latta & Howell, 2015).
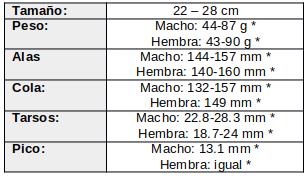
Descripción según los autores (Cleere, 1998:185; de La Peña 2020:227; Narosky & Yzurieta, 2010:194).
El macho es de color grisáceo-castaño teñido de rojizo, con una parte central de la corona claramente veteada de gris parduzco. El manto, espalda, rabadilla y coberteras supracaudales son gris ocráceo teñidas de castaño rojizo barradas y moteadas de color pardo.
Sus alas son anchas y redondeadas, con coberteras menores castañas, moteadas de marrón oscuro. Las coberteras alares primarias son pardas oscuras, mientras que el resto de las coberteras alares son grisáceas-marrón o parduzcas, moteadas y vermiculadas de beige y beige claro. Las escapulares son variables, con vexilos internos generalmente beige-parduzcos, castaños rojizos o castaños manchados de marrón, y vexilos externos grisáceos-parduzcos, con bordes y puntas beige. Primarias marrones con amplia banda blanca, en ángulo a través de P6-P10, que se extiende entre la mitad del ala y punta de P10 hasta la punta de P6. Tapadas primarias y secundarias marrones y ligeramente barradas de beige. Terciarias gris-parduzco teñidas de beige o castaño rojizo pálido y moteadas de marrón.
La cola es mas bien larga y redondeada, marrón, con R5-R3 con cantidades variables de blanco. R5 tiene blanco desde el raquis diagonalmente a través del vexilo interno hasta la punta; R4 es blanca en la mitad superior del vexilo externo y totalmente blanca sobre el vexilo interno; R3 generalmente es toda blanca, R2 abiertamente barrada de beige y el par central (R1) gris parduzco con tintes castaño-rojizos, barradas y moteadas de marrón. En reposo la cola es más larga que las alas (Hilty & Brown, 1986:294).
Loral y auriculares rojizas o castañas, moteadas de marrón. El anillo periocular beige y con raya blancuzca debajo del bigote. La barba y garganta son gris-parduzca, barrada con beige o castaño leonado. Diferentes autores, describen los ojos como reflectantes color rojo-rubí brillante (Snyder, 1966:126; Stiles & Skutch, 1989:200), pero también como naranja-rojizo en Trinidad y Panamá (ffrench, 1991:209; Wetmore, 1968:211; Ridgely & Gwynne 1989), dorado-rojizo en Texas (Oberholser, 1974:468), y rosado en Costa Rica (Slud, 1964:137)
El pico es marrón oscuro (Dickey & van Rossem, 1938:244) a marrón rosáceo con la punta negra (Oberholser, 1974:468). La base del pico es oscura y opaca. Interior de la boca rosado (Skutch, 1972:3). El iris es parduzco a marrón (Wetmore, 1968:209), marrón oscuro casi negro (Ridgway, 1914:538). El borde muy fino del párpado es marrón-amarillento opaco (Wetmore, 1968:209). Los lados del cuello beige, barrados de gris parduzco. Pequeña mancha blanca a cada lado de la parte inferior de la garganta, o parche blanco a través del total de la parte inferior de la garganta, con las plumas inferiores con puntas grisáceas parduzcas con una banda beige subterminal.
El pecho es grisáceo parduzco con tintes rojizos, moteado y vermiculado de marrón. El vientre, flancos, coberteras subcaudales y alares son ocráceas vermiculados de pardo oscuro.
Los tarsos y pies desnudos, ligeramente castaño o grisáceos rosados, con pies marrón oscuros y garras marrón opaco (Oberholser, 1974:468). Es un ave ágil y veloz, con tarsos más largos y músculos de las piernas más grandes que otros caprimúlgidos (Hartman, 1961:11).
La hembra es similar al macho, pero con una banda blanca más fina a través de P10-P8, ocasionalmente hasta P7, bordeada de beige y castaño rojizo desleído o beige sobre los vexilos externos. R4 y ocasionalmente R3 tienen las puntas apenas blancas, sobre los vexilos internos. El anillo periocular y la raya debajo del bigote son similares al macho. El iris es de color marrón oscuro casi negro. Los tarsos y pies desnudos son ligeramente castaño o grisáceos rosados, con pies marrón oscuros y garras marrón opaco.
En todas las subespecies los machos son más grandes que las hembras en todas las subespecies.
Existen variaciones geográficas que son enmascaradas por una considerable variación individual, así como por el plumaje dicromático, de marrón a castaño rojizo, a otro marcadamente gris. El plumaje también varía según el sexo y la altitud. En general, las poblaciones del norte son más pálidas y grisáceas y las del sur más oscuras y rojizas. Los plumajes oscuros se asocian con entornos más húmedos (Griscom, 1932:190). El tamaño varía desde tamaños más grandes en las poblaciones más al norte y sur, a más pequeño en las poblaciones de zonas intermedias (Ridgway, 1914:535).
Se reconocen seis subespecies, siguiendo a (Peters, 1940:192), que se distinguen fundamentalmente por las diferencias en plumaje y tamaño (Ridgway, 1914:535).
N. a. merrilli(Sennett, 1888). Residente, desde el valle del Río Grande inferior y al sur en la costa de Texas sobre el Golfo, hasta noreste de México en centro de Tamaulipas; Registrado en Veracruz y Puebla (Peters, 1940:192), donde tal vez sea visitante invernal (Ridgway, 1914:546).
Plumaje considerablemente gris con morfología castaño rojiza ausente o muy rara (Pyle, 1997, citado por Latta & Howell, 2020).
Tamaño promedio grande (ala macho > 172 mm, cola > 168 mm) (Latta & Howell, 2020).
N. a. yucatanensis (Nelson, 1901). Incluye N. a. nelsoni (Ridgway, 1912:91), y N. a. sumichrasti (Ridgway, 1912:91; Binford, 1989:136). Residente en este y oeste de México, desde el sur de Sinaloa a lo largo de la costa del Pacífico y desde Tamaulipas hacia el sur a lo largo de la costa del golfo, incluyendo la península de Yucatán, hasta norte y centro de Guatemala y Belice; también en Islas Mujeres y Cozumel (Peters, 1940:193). (ala macho < 176 mm, cola < 166 mm) (Latta & Howell, 2020).
N. a. insularis(Nelson, 1898). Residente en Islas Tres Marías (I. María Madre) (Peters, 1940:192).
Como el N. a. merrilli, pero más castaño en general y con cola corta (< 168 mm) (Latta & Howell, 2015)
N. a. albicollis (Gmelin, 1789). Incluye Caprimulgus guianensis (Gmelin, 1789), N. grallarius (Bonaparte, 1850), C. laticaudatus (Drapiez, 1851), N. a. obscurus (Cory, 1915) y N. a. intercedens (Griscom, 1929) (Monroe, 1968:360). Residente desde el oeste de Guatemala y Honduras hacia el sur, hasta este y sur de Venezuela, Trinidad, Las Guayanas hasta Perú, y norte y noreste de Brasil (Peters, 1940:193). También, probablemente en el este y sur de Colombia y sur de Ecuador hasta el este de Perú y norte de Bolivia
Similar al N. A. yucatanensis, con plumaje menos gris, más oscuro y castaño rojizo; ampliamente barrado y en promedio más pequeño (Latta & Howell, 2020).
N. a. gilvus (Bangs, 1902). Residente, en el litoral este de Panamá y en el norte de Colombia (desde el Río Sinú hacia el este, hasta Santa Marta); tal vez esté presente en el oeste de Venezuela (Peters, 1940:193).
Similar al N. a. albicollis, pero en general más pálido y blanquecino (menos rojizo); estrechamente barrado (Todd & Carriker, 1922:218; Wetmore, 1968:209; Latta & Howell, 2020).
N. a. derbyanus (Gould, 1838). Residente, desde centro de Brasil hacia el sur ( Matto Grosso, Goyaz, oeste Minas Geraes, Río Grande do Sul y Sao Paulo), a través del este de Bolivia, Paraguay, y noreste de Argentina (Misiones hasta Entre Ríos) (Peters, 1940:193; Latta & Howell, 2020).
NOTA TAXONÓMICA
El Nyctidromus albicollis ha sido clasificado, desde siempre, como el único miembro de un género monotípico, pero a través de estudios moleculares (Han et al., 2010:448), se ha encontrado que es hermano del Caprimulgus anthonyi, por lo que esta especie ha sido también colocada en el género Nyctidromus (Remsen et al., 2014: propuesta 12 – SACC: proponen incluir Caprimulgus anthonyi en Nyctidromus y Sigurdsson & Cracraft (2014:540) confirman la reasignación de C. anthonyi a Nyctidromus.
Sigurdsson & Cracraft (2014:541), presentan una nueva filogenia molecular de los Caprimulgidae (restringida al nuevo mundo) basándose en marcadores nucleares y mitocondriales de muestras obtenidas de ADN antiguo (muestras de pieles antiguas de museos). Con el apoyo de estos estudios genéticos y en comparaciones con trabajos previos, recomiendan las siguientes modificaciones taxonómicas en el caso del Nyctidromus albicollis (Gmelin, 1789).
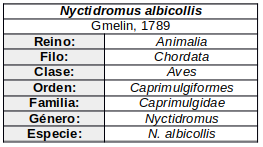
Nyctidromus (Gould, 1838) Especie:Nyctidromus albicollis (Gmelin, 1789).
Las evidencias genéticas sostienen la hipótesis que Caprimulgus anthonyi, C. hirundinaceus, y C. nigrescens están más estrechamente relacionados al Nyctidromus albicollis que otras especies del Nuevo Mundo previamente colocadas en el género Caprimulgus. Los resultados para C. anthonyi y C. nigrescens son coherentes con los de Han et al. (2010:448), pero ésta es la primera evidencia genética que sostiene este posicionamiento en el género Caprimulgus. Por lo tanto, se sugiere que estas tres especies deberían ser incluídas dentro del género Nyctidromus.
Éstas evidencias también respaldan la división las especies politípicas N. albicollis en tres especies filogenéticas diferentes: N. albicollis, N. derbyanus, y N. merrilli.
Especie: Nyctidromus derbyanus (Gould, 1838)
Hasta ahora Nyctidromus albicollis derbyanus. Es ascendida a la categoría de especie plena, hermana de la especie N. albicollis.
Notoriamente más grande que N. albicollis y N. merrilli. Espalda, corona, primarias externas y secundarias claaramente rojizas. No tan oscuras como en el N. albicollis.
Las evidencias obtenidas en estos estudios genéticos son consistentes con que el N. albicollis debe ser separado en tres especies filogenéticas. Los miembros de las subespecies nominales permanecen como N. albicollis, mientras que el N. a. derbyanus es ascendido al estado de especie bajo el nombre de N. derbyanus. Algunos individuos identificados como N. a. albicollis (especialmente al sur del Río Amazonas) podrían pertenecer a esta filoespecie.
Especie:Nyctidromus merrilli (Sennet, 1888)
Previamente N. a. merrilli, que incluye individuos asignados como N. a. insularis, N. a. intercedens, y N. a. yucatanensis. Es ascendido a estado de especie plena y es especie hermana del clado que contiene al N. albicollis y N. derbyanus.
El Nyctidromus merrilli es de tamaño intermedio, con plumaje más claro y gris que el N. albicollis y N. derbyanus, siendo notoria la coloración gris-pizarra de la corona y espalda. Destacan también los evidentes patrones negros de las cubiertas alares, que tanto en el N. albicollis como en el N. derbyanus tienden a ser marrones.
Es el guardacaminos más común. Su nombre Nyctidromus deriva de las raíces griegas nukti = nocturno y dromos = corredor. Su epíteto albicollis significa de cuello blanco y deriva de las raíces latinas albus = blanco y collis = cuello.
Se encuentra cerca de bosques o selvas en galería en óptimo estado de conservación, donde pasa el día camuflado en el suelo.
La mayoría de los registros se han producido en caminos vecinales, bosques secundarios y abiertos, o chacras en los límites de los parques. Donde hay rutas, el Curiango y otros caprimúlgidos son frecuentemente atropellados por vehículos.
Es el único de los atajacaminos completamente residente en EEUU, aunque su distribución está limitada al sur de Texas. Además, es el más común a través de su rango en México, y América Central y del Sur.
La actividad vocal del Curiango (la manera más fácil de detectarlo) es muy acentuada en la época estival o reproductiva, siendo muy infrecuente fuera de esta temporada.
NOMBRES COMUNES: Curiango (Argentina, Paraguay y Uruguay); Atajacaminos o dormilón de collar blanco (Argentina); Bujío, Gallinaciega común, Guardacaminos común (Colombia); Tapacaminos común (Costa Rica); Pucuyo coliblanco (Honduras); Chotacabras pauraque, Tapacaminos picuyo (México); Pocoyo tapacaminos (Nicaragua); Chotacabras común (Perú, Colombia); Aguaitacamino común (Venezuela); Bacurau, Curiango, Ju-jau, Kamaiurá y Aku-kúfjuruna en el Mato Grosso (Brasil); En guaraní: «Ibiyaú, urú-embudo, gïrá-ateí, cuñapirú» (Olrog, 1959:155; Chebez, 1996:134; Sick, 1997:418; de la Peña, 1999:90; de la Peña, 2020:227 )