Pasa la mayor parte del día buscando carroña, volando a alturas moderadas. Localizan la carroña con el olfato y la vista (Applegate, 1990:141), (Bang & Cobb, 1968:58); según Bang (1972:153), el bulbo olfatorio es más grande cuando se compara con los hemisferios cerebrales del C. atratus. Se los ha registrado, de forma aislada, cazando peces y animales vivos, generalmente por estar enfermos o discapacitados. Los estudios demuestran que comen carroña de presas más pequeñas que el C. atratus.
Se posa en árboles muertos, sin follaje, o en estructuras artificiales como torres de agua (Clark & Schmitt, 2017:92-93). Suelen hacerlo en grandes grupos, aunque durante el día buscan alimentos de forma independiente.
Pasa la noche en grupos comunitarios (pudiendo agruparse varios cientos de buitres para pasar la noche), a veces incluso con Jotes Cabeza Negra (Coragyps atratus) (Davis, 1979:124); (Buhnerkempe & Westemeier, 1984:496; (Clark & Schmitt, 2017:92-93)
A pesar de que anida en cuevas, no entra en ellas para descansar, excepto durante la época de reproducción. Durante la noche, reduce su temperatura corporal cerca de 6 grados centígrados hasta 34º C, volviéndose ligeramente hipotérmico. (Hatch, 1970:111); (Heath, 1962:234)
Frecuentemente posa con las alas extendidas. Se cree que esta postura cumple con múltiples funciones: secar las alas, calentar el cuerpo y combatir bacterias, sobre todo después de noches lluviosas o húmedas (Clark & Ohmart, 1985:353). El mismo comportamiento es observado en otros buitres americanos, buitres del Viejo Mundo, y cigüeñas. A menudo defeca sobre sus propias patas como medio para disipar el calor tras la evaporación del líquido contenido en las heces y la orina, un proceso conocido como urohidrosis (Hatch, 1970:111) De esta forma enfría los vasos sanguíneos que pasan por las patas y pies y causa manchas blancas por el ácido úrico que cubre las piernas. Cuando se encuentran expuestos a alta temperatura ambiente, controlan su temperatura extendiendo el cuello, desplegando las alas y mediante la urohidrosis (Arad & Bernstein, 1988:917); Arad et al. (1989:505), demostraron la extensa vascularización de la piel en la cabeza implume, cuello, nuca y pies; significa que los C. aura tienen un muy bien desarrollado sistema arterio-venoso intercambiador de calor en la cabeza, alas y patas (Arad et al., 1989:514)
Requiere de un gran esfuerzo para alzar el vuelo, batiendo sus alas mientras salta. El vuelo activo es lento, con aleteos profundos (cuando inclinan las alas hacia abajo, algunas veces tocan las puntas de las alas (Clark & Schmitt, 2017:92-93); (Mueller, 1972:221). Se elevan y planean con las alas en “V” (diedro). Las alas largas y anchas, y sus colas relativamente cortas están bien adaptadas para el vuelo planeado sobre la horizontal, balanceándose frecuentemente de un lado a otro (De la Peña & Salvador, 2016:151); (Narosky & Yzurieta, 2010:90); es un ejemplo de vuelo estático, aprovechando térmicas para mantener su altitud, a veces durante horas, sin necesidad de batir sus alas. Se desplaza en el suelo, con saltos torpes y muy desgarbados.
El C. aura tiene pocos depredadores naturales. Pueden caer presa de águilas reales, águilas, y búhos cornudos. Los huevos y polluelos pueden caer presa de mamíferos tales como mapaches y zorros. Su principal forma de defensa es regurgitar carne semi-digerida, una sustancia maloliente que irrita los ojos, con la que logra disuadir a la mayoría de las criaturas con intenciones de asaltar un nido de buitres. Un buitre también puede verse obligado a regurgitar una comida semi-digerida para deshacerse rápidamente del peso en su buche y poder tomar vuelo con el fin de huir de un depredador potencial. (Forbush, 1927: 91); (Vogel, 1959:213). No defienden su territorio.
VOCALIZACIONES
Como la mayoría de los buitres, carecen de complejidad en las vocalizaciones. Producen gruñidos y silbidos principalmente para disuadir a los depredadores.
Pemberton (1925:38) las describe así, “Emiten una voz distintiva nasal, un quejido de tono bajo, suave y repetido a intervalos de 3 segundos y que semeja a los quejidos de los cachorritos”; el autor hace notar que no abren el pico cuando emiten sonidos.
Según Allard (1934:515), antes de emprender vuelo emiten varias notas peculiares, comenzando con un corto, «tschuck, tschuck», repetido varias veces y seguido una nota quejosa que repite 2 á 3 veces. A continuación despegan y vuelan.
ALIMENTACIÓN
Busca alimento utilizando su sentido del olfato, un sentido compartido con los demás buitres del género Cathartes, pero poco común en el mundo de las aves. Localiza carroña al detectar el olor del etilmercaptano, un gas producido al inicio de la descomposición en los animales muertos. El lóbulo olfativo de su cerebro, responsable del procesamiento de los olores, es particularmente grande en comparación con otros animales. Se sabe que el Cathartes aura posee un sistema olfatorio anatómicamente bien desarrollado y fisológicamente funcional. Estudios anatómicos comparativos indican que, el tamaño relativo del bulbo olfatorio del C. aura es ocho veces más grande que el de 108 especies estudiadas por Bang & Cobb (1968:58); (Smith & Paselk, 1986:586)
Los seres humanos también aprovechan del sentido del olfato de los buitres americanos: se inyecta etilmercaptano en los oleoductos, y a continuación los ingenieros en busca de fugas monitorizan los vuelos de los buitres que buscan alimento.
Con su sentido del olfato superior tiene ventajas en la localización de la carroña y suele ser el primero en llegar a ella (Houston, 1986:318); (Houston, 1988:130); (Stager, 1964:3). Sin embargo, a pesar de su superior habilidad para detectar comida, los C. aura tienen pocas oportunidades de consumir los cadáveres de animales que descubren porque, los C. atratus frecuentemente se apropian de la comida. Las diferentes estrategias de alimentación de las dos especies se reflejan perfectamente en la altura a la que vuelan. El C. aura vuela cerca del suelo, donde la percepción olfatoria es más fuerte y sus posibilidades de encontrar cadáveres pequeños es mayor, mientras que el C. atratus busca comida volando más alto u observando los individuos que descienden, como señal para localizar las carcasas (Buckley et al., 1996:485); (Wallace et al.,1987:293); (Lemon, 1991:700).
Los C. aura pueden desgarrar carcasas de pequeño y mediano tamaño, pero no pueden con las de gran tamaño. Si ellos consiguen llegar a las carcasas pequeñas antes que otras especies, pueden obtener alimento sin interactuar con otros especímenes más dominantes que llegan más tarde. Aunque los C. aura individualmente son dominantes sobre un solo C. atratus, son intimidados los grupos de C. atratus, que a menudo suelen ser muy numerosos (50 individuos) (Wallace et al., 1987:293)
Es muy raro que los C. aura compartan la comida entre ellos y son reticentes a comer cerca de otros individuos (< 2m). Alimentarse en grandes grupos parece ser menos eficiente en los C. aura. Parece que defienden áreas a distancias mínimas unos de otros, en los sitios de alimentación. Las distancias individuales podrían ser mantenidas a través del reconocimiento pasivo del estatus social, reivindicando su posición dominante con picoteos amenazantes y más raramente con peleas. Estos actos o demostraciones agresivas pueden interferir con la alimentación del agresor (Prior et al., 1991:1555).
Se alimenta principalmente de una amplia variedad de carroña, desde pequeños mamíferos a herbívoros grandes (Coleman & Fraser, 1987:733); (de la Peña & Salvador, 2016:151); (Di Giacomo, 2005); (Salvador et al., 2017:103); (Hiraldo et al., 1991:321); (Prior, 1990:709), prefiriendo los recién muertos (Lemon, 1991:702), y evitando los cadáveres que han llegado al punto de putrefacción (Owre & Northington,1961:200). Raramente se alimenta de materia vegetal (vegetación de ribera, calabazas u otros cultivos), y de insectos vivos y otros invertebrados (Crafts Jr., 1968:327); (Paterson, 1984:469). (Pinto (1965:276), lo observó alimentándose de frutos de Palmera (Elaeis guineensis)
Se ha observado a los C. aura y C. atratus consumiendo huevos de tortugas verdes (Chelonia mydas) (Fowler, 1979:946) y a los C. aura alimentándose de huevos en colonias de aves marinas (Kirk and Mossman, 2020), de cocodrilo americano (Crocodylus acutus) (Platt et al., 2014:39) y de phoenicopterus chilensis (Sosa, 1999:95)
Es poco común que cace y mate a sus presas (Galding & Glading, 1970:244); (Mueller & Berger, 1967:431). Hamilton (1941:254), comenta que el ataque de Cathartes aura a unas crías de cerdos, cerca de Fort Myers (Florida), pudo haber sido un “caso de error de identificación” (Parmalee, 1954:443). A menudo se le puede observar a lo largo de las carreteras (Rea, 1973:210), alimentándose de los cadáveres de animales atropellados, o cerca del agua, alimentándose de peces varados en bancos a orillas de ríos (Jackson et al, 1978:142); (Platt et al., 2009:840). Al igual que otros buitres, desempeña un papel importante en su ecosistema, librando al ambiente de la carroña que de otro modo podría producir enfermedades.
Los autores (Coleman et al.,1985:291), mientras estudiaban los hábitos alimenticios de los Cathartes, los observaron comiendo sal a 4 km al sur de Gettysburg (Pennsylvania).
REPRODUCCIÓN
A menudo, el apareamiento entre los C. aura, es precedido por una especie de danza gregaria: varios individuos se juntan en el suelo de un claro, rodeando a una hembra; realizan movimientos de salto con las alas extendidas en todo el perímetro del círculo; un individuo salta hacia su vecino, el cual a continuación salta y se aproxima a un tercero, etc, mientras, otros buitres posados en árboles cercanos bajan para sumarse a la “danza”, mientras otros “bailarines” se retiran (Loftin & Tyson, 1965:193). También realizan vuelos de cortejo, lanzándose o persiguiéndose entre sí.
No construyen nido. Depositan los huevos en cuevas pequeñas, sobre el piso, bajo vegetación espesa, e incluso en edificios y coches abandonados (Igl & Peterson, 2010:73). Tiene más cuidado en la elección del sitio que en la construcción del mismo (Coles, 1944:222)
El Cathartes aura septentrionalis, por el contrario, tiene una mayor diversificación en la elección de sitios para anidar. Se han encontrado nidos en acantilados, así como, en pantanos, cuevas, laderas abruptas o escondidos en ciénagas saladas. Ubica sus nidos en bosques, arbustos y matorrales, a lo largo de cursos de agua y en estructuras abandonadas (Coles, 1944:219); (Sarasola, 2000:60)
Moroni et al. (2014:51-52), comenta: “El Jote Cabeza Roja nidifica en forma variada en nuestro país: en huecos de árboles (Hartert & Venturi 1909, de la Peña 2005), entre rocas (Sarasola et al. 2000), en el suelo debajo de arbustos (De Lucca et al. 2012), en la base de matas de paja y en ocasiones en cuevas (Woods 1988)”.
Se reproducen una vez al año. El inicio de la temporada reproductiva varía dependiendo de la latitud en que permanece. En el norte del área de distribución se inicia entre principios de marzo hasta junio
Las hembras suelen poner dos huevos (con diferencia de 1 día), aunque la puesta puede variar entre uno y tres huevos. Huevos ovoides, blanquecinos, blancos grisáceos o verdosos con manchas castañas oscuras y pintas grises, más concentradas en el polo mayor. Algunos con pocas pintas y manchas. (Coles, 1944:223); (De la Peña, 2013:87)
La incubación se inicia una vez que está completa la puesta. Ambos padres participan en la incubación, que dura entre 30-40 días (Coles 1944:227). Los periodos de incubación más cortos reportados son de 32 días (Gentry, 1882) y Audubon, 1835); 30 días (Burns, 1937), (Jackson, 1903) y (Kempton, 1927); (Coles, 1944:224).
Crías altriciales, nacen ciegos, desnudos e indefensos. Durante unas 10 u 11 semanas, cuidan de los polluelos y les dan de comer regurgitando. Cuando los adultos están amenazados mientras anidan, pueden regurgitar sobre el intruso, fingir la muerte, o huir del nido. Si los polluelos se ven amenazados en el nido, se defienden silbando y regurgitando. Los polluelos permanecen en el nido durante 9 a 10 semanas. Los grupos familiares permanecen juntos hasta el otoño.
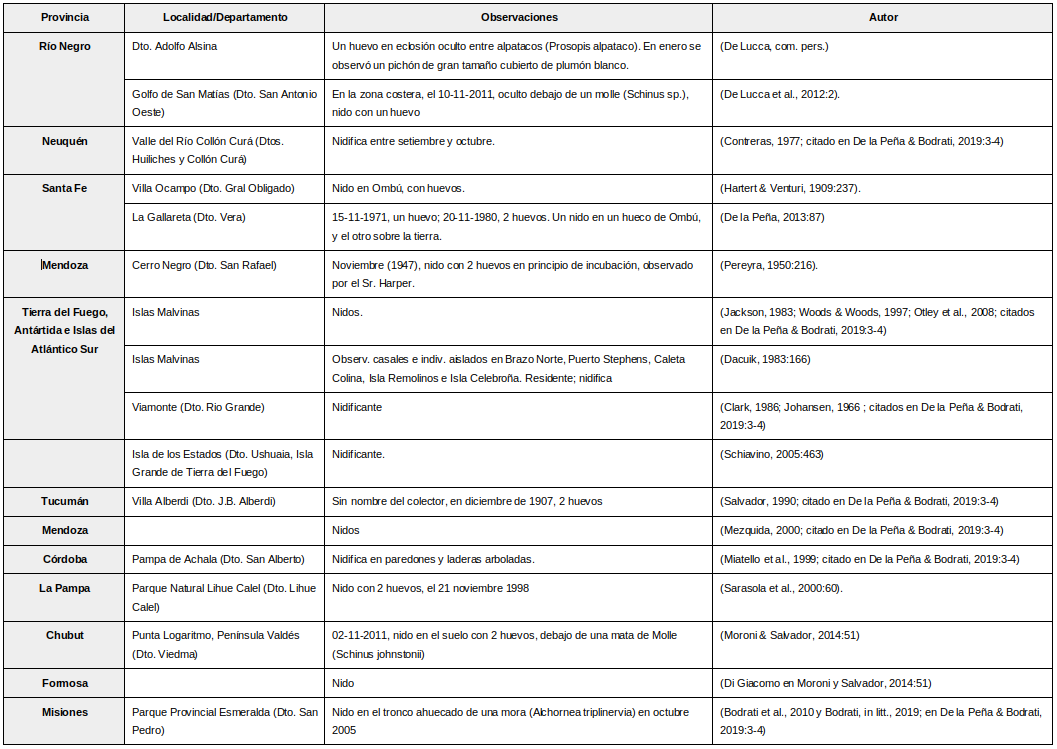
Argentina: Aunque es una especie ampliamente distribuida en el continente americano (Bendire, 1892:163), su rango de reproducción en Argentina no es bien conocido. Se localizaron nidos en Mendoza, Santa Fe, Tucumán, Formosa, Misiones, La Pampa, Córdoba, Neuquén, Río Negro, Chubut, Tierra del Fuego e Islas Malvinas, en los meses, octubre, noviembre y diciembre (De la Peña & Salvador, 2016:151); (De la Peña, 2013:87); (De la Peña & Bodrati, 2019:3-4)
Se citan los registros de nidificación conocidos: